
De fettsyror är organiska makromolekyler härledda från kolväten, som består av långa kedjor av kol- och väteatomer som är hydrofoba (de är fettlösliga) och är den strukturella grunden för fetter och lipider.
De är mycket olika molekyler som skiljer sig från varandra genom längden på deras kolvätekedjor och närvaron, antalet, positionen och / eller konfigurationen av deras dubbelbindningar..
I lipider från djur, växter, svampar och mikroorganismer såsom bakterier och jäst har mer än 100 olika klasser av fettsyror beskrivits och anses vara arter och vävnadsspecifika i de flesta levande varelser.
Oljorna och fetterna som människan konsumerar dagligen är av animaliskt eller vegetabiliskt ursprung, de består huvudsakligen av fettsyror.
Artikelindex

Fettsyramolekyler utför viktiga funktioner på mobilnivå, vilket gör dem till väsentliga komponenter och eftersom vissa av dem inte kan syntetiseras av djur, måste de få dem från kosten.
Fettsyror är ovanliga som fria arter i cellcytosolen, så de finns i allmänhet som en del av andra molekylära konjugat, såsom:
- Lipider, i biologiska membran.
- Triglycerider eller fettsyraestrar, som fungerar som reserver i växter och djur.
- Vaxar, som är fasta estrar av långkedjiga fettsyror och alkoholer.
- Andra liknande ämnen.
Hos djur lagras fettsyror i cellernas cytoplasma som små droppar fett sammansatta av ett komplex som kallas triacylglycerol, vilket är inget annat än en glycerolmolekyl till vilken den, i var och en av dess kolatomer, har en fettsyrakedja. av esterlänkar.
Medan bakterier har korta och vanligtvis enkelomättade fettsyror, är det i naturen vanligt att hitta fettsyror vars kedjor har jämnt antal kolatomer, vanligtvis mellan 14 och 24, mättade, enkelomättade eller fleromättade..
Fettsyror är amfipatiska molekyler, det vill säga de har två kemiskt definierade regioner: en hydrofil polär region och en hydrofob apolär region..
Den hydrofoba regionen består av en lång kolvätekedja som i kemiska termer inte är särskilt reaktiv. Den hydrofila regionen å andra sidan består av en terminal karboxylgrupp (-COOH), som beter sig som en syra.
Denna terminala karboxylgrupp eller karboxylsyra joniseras i lösning, är mycket reaktiv (kemiskt sett) och är mycket hydrofil och representerar således ett kovalent bindningsställe mellan fettsyran och andra molekyler..
Längden på fettsyrornas kolvätekedjor har vanligtvis jämnt antal kolatomer, och detta är nära relaterat till den biosyntetiska processen genom vilken de produceras, eftersom deras tillväxt sker i par kol..
De vanligaste fettsyrorna har kedjor med mellan 16 och 18 kolatomer och hos djur är dessa kedjor inte grenade.
Fettsyror klassificeras i två stora grupper efter karaktären av bindningarna som komponerar dem, det vill säga enligt närvaron av enkelbindningar eller dubbelbindningar mellan kolatomerna i deras kolvätekedjor.
Således finns det mättade och omättade fettsyror.
- Mättade fettsyror har endast enstaka kol-kolbindningar och alla deras kolatomer är "mättade" eller bundna till vätemolekyler..
- Omättade fettsyror har en eller flera kol-kol-dubbelbindningar och inte alla dessa är bundna till en väteatom.
Omättade fettsyror delas också upp efter antalet omättningar (dubbelbindningar) i enkelomättade, de med endast en dubbelbindning och fleromättade, de med mer än en.
De har vanligtvis mellan 4 och 26 kolatomer kopplade genom enstaka bindningar. Dess smältpunkt är direkt proportionell mot kedjans längd, det vill säga dess molekylvikt.
Fettsyror som har mellan 4 och 8 kol är flytande vid 25 ° C och är de som utgör ätliga oljor, medan de med mer än 10 kolatomer är fasta.
Bland de vanligaste är laurinsyra, som finns rikligt med palm- och kokosolja; palmitinsyra, finns i palm, kakao och ister och stearinsyra, finns i kakao och hydrerade oljor.
De är fettsyror med mycket mer stabilitet än omättade fettsyror, särskilt mot oxidation, åtminstone under fysiologiska förhållanden.
Eftersom enstaka kol-kolbindningar kan rotera fritt är mättade fettsyror mycket flexibla molekyler, även om steriskt hinder gör den helt utsträckta strukturen den mest energiskt stabila..
Dessa fettsyror är mycket reaktiva och utsatta för mättnad och oxidation. De är vanliga i växter och marina organismer. De med endast en dubbelbindning är kända som enkelomättade eller monoenoiska, medan de med mer än två är kända som fler- eller fleromättade..
Närvaron av dubbelbindningar är vanligt mellan kolatomerna mellan positionerna 9 och 10, men detta betyder inte att enomättade fettsyror med en omättnad i en annan position inte hittas.
Till skillnad från mättade är omättade fettsyror listade inte från den terminala karboxylgruppen utan enligt positionen för den första C-C-dubbelbindningen, så att de är uppdelade i två grupper, omega-6- eller ~ 6-syror och omega-3. eller ω3.
Omega-6-syror har den första dubbelbindningen vid kolnummer 6 och omega-3-syror har den vid kolnummer 3. Namnet ω ges av dubbelbindningen närmast den slutliga metylgruppen.
Dubbelbindningar finns också i två geometriska konfigurationer som kallas "cis " Y "trans".
De flesta av de naturliga omättade fettsyrorna har konfiguration "Cis”Och de dubbla bindningarna av fettsyror som finns i kommersiella (hydrerade) fetter finns i "trans".
I fleromättade fettsyror separeras vanligtvis två dubbelbindningar från varandra med minst en metylgrupp, det vill säga en kolatom bunden till två väteatomer..
Fettsyror har flera funktioner i levande organismer och, som nämnts ovan, är en av deras väsentliga funktioner som en väsentlig del av lipider, som är huvudkomponenterna i biologiska membran och en av de tre mest förekommande biomolekylerna i organismer. Lever tillsammans med protein och kolhydrater.
De är också utmärkta energisubstrat tack vare vilka stora mängder energi erhålls i form av ATP och andra mellanliggande metaboliter..
Med tanke på att djur till exempel inte kan lagra kolhydrater utgör fettsyror den huvudsakliga energilagringskällan som kommer från oxidationen av socker som konsumeras i överskott..
Mättade kortkedjiga fettsyror i tjocktarmen deltar i att stimulera absorptionen av vatten och natrium-, klorid- och bikarbonatjoner; Dessutom har de funktioner vid produktion av slem, i spridning av kolonocyter (kolonceller) etc..
Omättade fettsyror är särskilt rikliga i ätbara vegetabiliska oljor, som är viktiga för alla människors kost.

Andra deltar som ligander av vissa proteiner med enzymatiska aktiviteter, så de är viktiga med avseende på deras effekter på energimetabolismen i cellerna där de finns..
Nedbrytningen av fettsyror är känd som β-oxidation och inträffar i mitokondrier i eukaryota celler. Tvärtom förekommer biosyntes i cytosolen hos djurceller och i kloroplasterna (fotosyntetiska organeller) i växtceller..
Det är en process som är beroende av acetyl-CoA, malonyl-CoA och NADPH, den förekommer i alla levande organismer och hos "högre" djur såsom däggdjur. Det är till exempel mycket viktigt i lever- och fettvävnader, liksom i bröstkörtlarna..
NADPH som används för denna väg är huvudsakligen en produkt av de NADP-beroende oxidationsreaktionerna av pentosfosfatvägen, medan acetyl-CoA kan komma från olika källor, till exempel från oxidativ dekarboxylering av pyruvat, från Krebs-cykeln och β-oxidation av fettsyror.
Biosyntesvägen, som den för β-oxidation, regleras starkt i alla celler av allosteriska effektorer och kovalenta modifieringar av enzymer som deltar i reglering..
Vägen börjar med bildandet av en metabolisk mellanprodukt känd som malonyl-CoA från en acetyl-CoA-molekyl och katalyseras av ett multifunktionellt enzym som kallas acetyl-CoA-karboxylas..
Denna reaktion är en additionsreaktion av en biotinberoende karboxylmolekyl (-COOH, karboxylering) och sker i två steg:
Hos djur sker bildandet av kolhydratkedjor av fettsyror vidare genom sekventiella kondensationsreaktioner katalyserade av ett multimert och multifunktionellt enzym som kallas fettsyrasyntas..
Detta enzym katalyserar kondensationen av en acetyl-CoA-enhet och flera malonyl-CoA-molekyler som produceras från acetyl-CoA-karboxylasreaktionen, en process under vilken en molekyl CO2 frigörs för varje malonyl-CoA som det tillför.
De växande fettsyrorna förestras till ett protein som kallas "acylbärarprotein" eller ACP, som bildar tioestrar med acylgrupper. På E coli detta protein är en 10 kDa polypeptid, men hos djur är det en del av fettsyrasyntas-komplexet.
Brottet av dessa tioesterbindningar frigör stora mängder energi, vilket gör det möjligt, termodynamiskt sett, att kondenssteg förekommer i den biosyntetiska vägen..
I bakterier motsvarar fettsyrasyntasaktivitet faktiskt sex oberoende enzymer som använder acetyl-coA och malonyl-coA för att bilda fettsyror och med vilka sex olika enzymatiska aktiviteter är associerade.
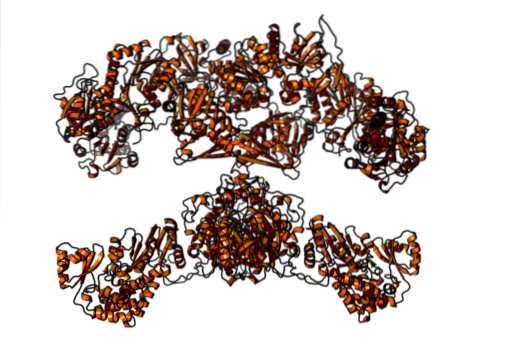
Hos däggdjur är fettsyrasyntas däremot ett multifunktionellt homodimert enzymkomplex med cirka 500 kDa molekylvikt, som har sex olika katalytiska aktiviteter och som acylbärarproteinet associerar.
Tiolgrupperna i cysteinresterna som ansvarar för bindningen av metaboliska mellanprodukter till ACP-enzymet måste fyllas med nödvändiga acylgrupper innan syntesen börjar..
För detta överförs acetylgruppen i acetyl-coA till tiolgruppen (-SH) i en av cysteinresterna i ACP-underenheten av fettsyrasyntas. Denna reaktion katalyseras av ACP-acyl-transferas-underenheten..
Acetylgruppen överförs sedan från ACP till en annan cysteinrest vid det katalytiska stället för en annan enzymunderenhet av komplexet känt som β-ketoacyl-ACP-syntas. Således "primas" enzymkomplexet för att börja syntes..
Malonyl-CoA som produceras av acetyl-CoA-karboxylas överförs till tiolgruppen i ACP och under denna reaktion förloras CoA-delen. Reaktionen katalyseras av malonyl-ACP-transferasunderenheten i fettsyrasyntaskomplexet, som sedan producerar malonyl-ACP.
Under denna process är malonylgruppen kopplad till ACP respektive β-ketoacyl-ACP-syntas genom en ester respektive en annan sulfhydrylbindning..
Enzymet p-ketoacyl-ACP-syntas katalyserar överföringen av acetylgruppen som var bunden till den i "priming" -steget till 2-kolet i malonylgruppen som i föregående steg överfördes till ACP.
Under denna reaktion frigörs en CO2-molekyl från malonyl, vilket motsvarar den CO2 som tillhandahålls av bikarbonat i karboxyleringsreaktionen av acetyl-CoA-karboxylas. Acetoacetyl-ACP produceras sedan.
Β-ketoacyl-ACP-reduktasunderenheten katalyserar den NADPH-beroende reduktionen av acetoacetyl-ACP och bildar därigenom D-β-hydroxibutyryl-ACP.
I detta steg bildas trans-a, P-acyl-ACP eller ~ 2-omättad-acyl-ACP (kratonyl-ACP), en produkt av dehydratiseringen av D-P-hydroxibutyryl-ACP genom inverkan av enoyl- underenhet. ACP-hydratas.
Senare reduceras kratonyl-ACP till butyryl-ACP genom en NADPH-beroende reaktion katalyserad av enoyl-ACP-reduktasunderenheten. Denna reaktion fullbordar den första av sju cykler som behövs för att producera palmitoyl-ACP, vilket är en föregångare till nästan alla fettsyror..
Butyrylgruppen överförs från ACP till tiolgruppen i en cysteinrest i β-ketoacyl-ACP-syntas, med vilken ACP kan acceptera en annan malonylgrupp från malonyl-CoA.
På detta sätt är reaktionen som inträffar kondensationen av malonyl-ACP med buturyl-β-ketoacyl-ACP-syntas, vilket ger upphov till β-ketohexanoyl-ACP + CO2.
Palmitoyl-ACP som uppstår från de efterföljande stegen (efter tillsats av ytterligare 5 malonylenheter) kan frisättas som fri palmitinsyra tack vare aktiviteten av tioesterasenzymet, den kan överföras till CoA eller införlivas i fosfatidsyra för fosfolipiden och triacylglyceridsyntesväg.
Fettsyrasyntaset hos de flesta organismer stannar vid syntesen av palmitoyl-ACP, eftersom det katalytiska stället för β-ketoacyl-ACP-syntas-subenheten har en konfiguration i vilken endast fettsyror av den längden kan rymmas.
Dessa är relativt vanliga i marina organismer och syntetiseras också av ett fettsyrasyntas-komplex. Emellertid sker "priming" -reaktionen med en längre molekyl, propionyl-ACP, med tre kolatomer..
Palmitinsyra, som diskuterat, fungerar som en föregångare för många mättade och omättade fettsyror med längre kedja. Processen med "förlängning" av fettsyror sker i mitokondrierna, medan införandet av omättningar sker i huvudsak i det endoplasmatiska retikulumet.
Många organismer omvandlar sina mättade fettsyror till omättade som en anpassning till låga omgivningstemperaturer, eftersom detta gör att de kan hålla smältpunkten för lipider under rumstemperatur..
Många av egenskaperna hos fettsyror beror på deras kedjelängd och närvaron och antalet omättningar:
- Omättade fettsyror har lägre smältpunkter än mättade fettsyror med samma längd.
- Längden av fettsyror (antalet kolatomer) är omvänt proportionell mot molekylens fluiditet eller flexibilitet, det vill säga de "kortare" molekylerna är mer flytande och vice versa..
I allmänhet består flytande fettsubstanser av kortkedjiga fettsyror med närvaro av omättningar.
Växter har stora mängder omättade fettsyror såväl som djur som lever vid mycket låga temperaturer, eftersom dessa, som komponenter i lipiderna i cellmembran, ger dem större flytbarhet under dessa förhållanden..
Under fysiologiska förhållanden orsakar närvaron av en dubbelbindning i kolvätekedjan av en fettsyra en krökning på cirka 30 °, vilket gör att dessa molekyler upptar ett större utrymme och minskar styrkan i deras van der Waals-interaktioner..
Närvaron av dubbelbindningar i fettsyror associerade med lipidmolekyler har direkta effekter på graden av "förpackning" som dessa kan ha i de membran till vilka de tillhör och har således också effekter på membranproteiner.
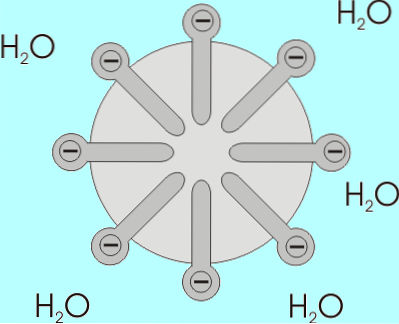
Lösligheten för fettsyror minskar när deras kedjelängd ökar, så de är omvänt proportionella. I vattenhaltiga och lipidblandningar associeras fettsyror med strukturer som kallas miceller..
En micell är en struktur där de alifatiska kedjorna av fettsyror är "inneslutna" och därmed "utvisar" alla vattenmolekyler och på vars yta karboxylgrupperna finns..
Nomenklaturen för fettsyror kan vara något komplex, speciellt om man hänvisar till de vanliga namnen de får, som ofta är relaterade till någon fysisk-kemisk egenskap, med platsen där de finns eller andra egenskaper.
Många författare anser att eftersom dessa molekyler joniseras vid fysiologiskt pH tack vare den terminala karboxylgruppen, bör man hänvisa till dem som "karboxylater" med användning av termineringen "ato ".
Enligt IUPAC-systemet görs uppräkningen av kolatomerna i en fettsyra från karboxylgruppen vid den polära änden av molekylen och de två första kolatomerna fästa vid denna grupp kallas α respektive β. Den terminala metylen i kedjan innehåller kolatomen ω.
I den systematiska nomenklaturen får de i allmänhet namnet "moder" kolväte (kolväten med samma antal kolatomer) och dess slut "eller" för "Oico", om det är en omättad fettsyra, läggs slutet till "Enoic".
Tänk till exempel fallet med en C18 (C18) fettsyra:
- Eftersom kolvätet med samma antal kolatomer är känt som oktadekan, kallas den mättade syran "oktadekansyra" Nåväl "oktadekanoat”Och dess vanliga namn är stearinsyra.
- Om du har en dubbelbindning mellan några par kolatomer i din struktur kallas det "oktadecensyra"
- Om den har två dubbelbindningar c - c kallas den "Octadecadienoic acid" och om du har tre "oktadekatriensyra".
Om du vill sammanfatta nomenklaturen används 18: 0 för 18-kolfettsyran och inga dubbelbindningar (mättad) och, beroende på graden av omättnad, skrivs 18: 1 istället för noll för en molekyl med en omättnad, 18: 2 för en med två omättningar och så vidare.
Om du vill ange mellan vilka kolatomer som är dubbelbindningarna i omättade fettsyror, använd symbolen ∆ med ett numeriskt överskrift som anger platsen för omättnad och prefixet "Cis"Eller "trans", beroende på konfigurationen av detta.


Ingen har kommenterat den här artikeln än.