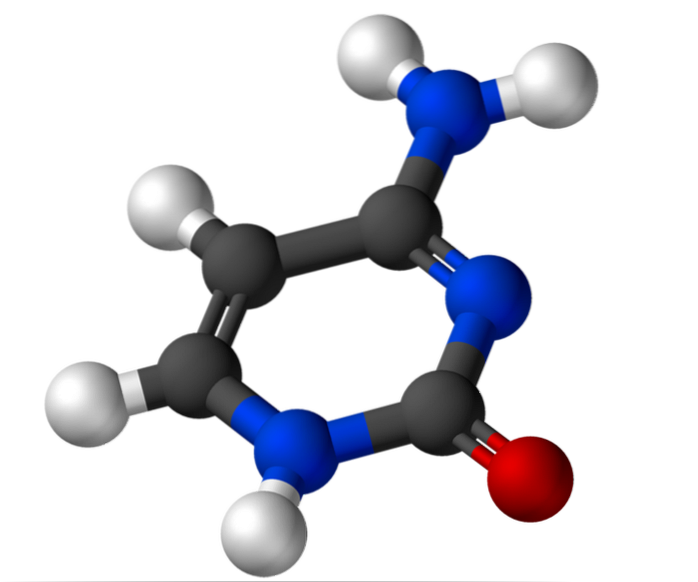
De cytosin Det är en kvävebas av pyrimidintyp som används för biosyntes av cytidin-5'-monofosfat och deoxicytidin-5'-monofosfat. Dessa föreningar tjänar till biosyntes av deoxiribonukleinsyra (DNA) respektive ribonukleinsyra (RNA). DNA lagrar genetisk information och RNA har olika funktioner.
I levande saker finns inte cytosin i fri form utan bildar ofta ribonukleotider eller deoxiribonukleotider. Båda typerna av föreningar har en fosfatgrupp, en ribos och en kvävebas..
2-kolet i ribos har en hydroxylgrupp (-OH) på ribonukleotider och en väteatom (-H) på deoxiribonukleotider. Beroende på antalet närvarande fosfatgrupper finns det cytidin-5'-monofosfat (CMP), cytidin-5'-difosfat (CDP) och cytidin-5'-trifosfat (CTP).
Deoxygenerade ekvivalenter kallas deoxycytidin-5'-monofosfat (dCMP), deoxycytidin-5'-difosfat (dCDP) och deoxycytidin-5'-trifosfat (dCTP)..
Cytosin, i dess olika former, deltar i olika funktioner, såsom DNA- och RNA-biosyntes, glykoproteinbiosyntes och reglering av genuttryck..
Artikelindex
Cytosin, 4-amino-2-hydroxipyrimidin, har den empiriska formeln C4H5N3O, vars molekylvikt är 111,10 g / mol, och renas som ett vitt pulver.
Strukturen av cytosin är en plan aromatisk heterocyklisk ring. Våglängden för maximal absorbans (ʎmax) är vid 260 nm. Smältemperaturen för cytosin överstiger 300 ° C.
För att bilda en nukleotid är cytosin bundet kovalent, genom kväve 1, via en N-beta-glykosidbindning till 1'-kolet i ribos. 5'-kolet förestras med en fosfatgrupp.
Pyrimidinnukleotidbiosyntes har en gemensam väg, bestående av sex enzymkatalyserade steg. Vägen börjar med karbamoylfosfatbiosyntes. I prokaryoter finns det bara ett enzym: karbamoylfosfatsyntas. Detta är ansvarigt för syntesen av pyrimidiner och glutamin. I eukaryoter finns karbamoylfosfatsyntas I och II, som är ansvariga för biosyntesen av glutamin och pyrimidiner.
Det andra steget består av bildandet av N-karbamoylaspartat, från karboylfosfat och aspartat, en reaktion katalyserad av aspartattranskabamoylas (ATCas).
Det tredje steget är syntesen av L-dihydrorotat, vilket orsakar stängningen av pyrimidinringen. Detta steg katalyseras av dihydrootas..
Det fjärde steget är bildandet av orotat, vilket är en redoxreaktion katalyserad av dihydroorotatdehydrogenas..
Det femte steget består av bildandet av orotidylat (OMP) med användning av fosforibosylpyrofosfat (PRPP) som substrat, och orotatfosforibosyltransferas som en katalysator..
Det sjätte steget är bildandet av uridylat (uridin-5'-monofosfat, UMP), en reaktion katalyserad av ett OMP-dekarboxylas..
Nästa steg består av fosforylering av UMP, katalyserad av kinaser, för att bilda UTP, och överföringen av en aminogrupp från glutamin till UTP för att bilda CTP, en reaktion katalyserad av CTP-syntetas..
Hos däggdjur sker reglering på nivån av karbamoylfosfatsyntas II, ett enzym som finns i cytosolen, medan karbamoylfosfatsyntas I är mitokondrie.
Karbamoylfosfatsyntas II regleras av negativ återkoppling. Dess regulatorer, UTP och PRPP, är respektive hämmare och aktivator av detta enzym.
I icke-levervävnader är karbamoylfosfatsyntas II den enda källan till karbamoylfosfat. I levern, under förhållanden med överskott av ammoniak, producerar karbamoylfosfatsyntas I i mitokondrierna karbamoylfosfat, som transporteras till cytosolen, varifrån det går in i pyrimidinbiosyntesvägen..
En annan regleringspunkt är OMP-dekarboxylas, som regleras av konkurrerande hämning. Dess reaktionsprodukt, UMP, konkurrerar med OMP om bindningsstället på OMP-dekarboxylas..
Återvinning av pyrimidiner har funktionen att återanvända pyrimidiner utan behov av de novo biosyntes och undvika den nedbrytande vägen. Återvinningsreaktionen katalyseras av pyrimimidinfosforibosyltransferas. Den allmänna reaktionen är som följer:
Pyrimidin + PRPP -> pyrimidinnukleosid 5'-monofosfat + PPi
Hos ryggradsdjur finns pyrimimidinfosforibosyltransferas i erytrocyter. Substratpyrimidinerna för detta enzym är uracil, tymin och orotat. Cytosin återvinns indirekt från uridin-5'-monofosfat.
Under DNA-replikering kopieras informationen i DNA: t till DNA: t med ett DNA-polymeras..
RNA-biosyntes kräver deoxinukleotidtrifosfat (dNTP), nämligen: deoxitymidintrifosfat (dTTP), deoxicytidintrifosfat (dCTP), deoxiadenintrifosfat (dATP) och deoxyguanintrifosfat (dGTP). Reaktionen är:
(DNA)n rester + dNTP -> (DNA)n + 1 rest + PPi
Hydrolys av oorganiskt pyrofosfat (PPi) ger energi för RNA-biosyntes.
I DNA-dubbelhelixen är en ensträngad purin kopplad till den motsatta strängen pyrimidin genom vätebindningar. Således är cytosin alltid kopplat till guanin genom tre vätebindningar: adenin är kopplat till tymin genom två vätebindningar..
Vätebindningar bryts när en lösning av renat nativt DNA, vid pH 7, utsätts för temperaturer över 80 ° C. Detta gör att DNA-dubbelhelixen bildar två separata strängar. Denna process är känd som denaturering..
Temperaturen vid vilken 50% av DNA denatureras är känd som smälttemperaturen (Tm). DNA-molekyler vars förhållande mellan guanin och cytosin är högre än det för tymin och adenin har högre Tm-värden än de vars basförhållande är invers.
Ovan beskrivna utgör det experimentella beviset för att ett större antal vätebindningar bättre stabiliserar de nativa DNA-molekylerna..
Nyligen konstaterades att DNA från kärnan i mänskliga celler kan anta interstruktiva motivstrukturer (iM). Dessa strukturer förekommer i regioner som är rika på cytosin.
IM-strukturen består av fyra DNA-strängar, till skillnad från klassiskt dubbelsträngat DNA som har två strängar. Mer specifikt är två parallella duplexkedjor isärlagda i en antiparallell orientering och hålls samman av ett par hemiprotonerade cytosiner (C: C+).
I det mänskliga genomet finns iM-strukturer i regioner som promotorer och telomerer. Antalet iM-strukturer är högre under G1 / S-fasen i cellcykeln, där transkriptionen är hög. Dessa regioner är proteinigenkänningsställen involverade i aktiveringen av transkriptionsmaskineriet..
Å andra sidan, i regionerna som är rika på på varandra följande guaninbaspar (C), tenderar DNA: t att anta A-helixformen under uttorkande förhållanden. Denna form är typisk för RNA och blandade DNA-RNA-dubbelband under transkription och replikering, och vid vissa tidpunkter när DNA är bundet till proteiner..
På varandra följande basregioner av cytosin har visat sig skapa ett elektropositivt plåster i huvuddelen av DNA. Därför antas dessa regioner binda till proteiner, vilket predisponerar vissa genomregioner för genetisk bräcklighet..
Under transkription kopieras informationen i DNA till RNA med ett RNA-polymeras. RNA-biosyntes kräver nukleosidtrifosfat (NTP), nämligen: cytidintrifosfat (CTP), uridintrifosfat (UTP), adenintrifosfat (ATP) och guanintrifosfat (GTP). Reaktionen är:
(RNA)n rester + NTP -> (RNA)n + 1 rest + PPi
Hydrolys av oorganiskt pyrofosfat (PPi) ger energi för RNA-biosyntes.
Den sekventiella överföringen av hexoser för att bilda oligosackarider, O-kopplade till proteiner, sker från nukleotidprekursorer.
Hos ryggradsdjur består det sista steget i O-länkad oligosackaridbiosyntes av tillsatsen av två sialinsyrarester (N-acetylneuramin) från en cytidin-5'-monofosfat (CMP) föregångare. Denna reaktion inträffar i transgolgisäcken.
Tetrahydrofolatsyra (FH4) är en källa till -CH-grupper3, och är nödvändig för biosyntes av dTMP från dUMP. Dessutom bildas FH2. Reduktionen av FH2 till FH4 kräver ett reduktas av folat och NADPH. Vissa folatreduktashämmare, såsom aminopterin och metotrexat, används vid cancerbehandlingar.
Metotrexan är en konkurrerande hämmare. Folatreduktas binder med 100 gånger mer affinitet till denna hämmare än till dess substrat. Aminopterin fungerar på liknande sätt.
Hämningen av folatreduktas hindrar indirekt biosyntesen av dTMP, och därför den av dCTP. Direkt inhibering sker av hämmare av tymidylatsyntetasenzymet, som katalyserar dTMP från dUMP. Dessa hämmare är 5-fluorouracil och 5-fluoro-2-deoxyuridin..
Till exempel är 5-fluoracyl inte i sig en hämmare utan omvandlas först, i återvinningsvägen, till deoxiuridinmfosfat d (FdUMP), som binder och hämmar tymidylatsyntetas..
Ämnen som är analoga med glutamin, azaserin och acivicin hämmar glutaminamidotransferas. Azarin var ett av de första ämnena som upptäcktes som fungerar som en självmordsinaktivator.


Ingen har kommenterat den här artikeln än.