De konvergerande evolution Det är framväxten av fenotypiska likheter i två eller flera linjer oberoende av varandra. Generellt observeras detta mönster när de involverade grupperna utsätts för liknande miljöer, mikromiljöer eller livsstilar som översätts till motsvarande selektiva tryck..
Således ökar de fysiologiska eller morfologiska egenskaperna i fråga den biologiska adekvaten (kondition) och konkurrenskraft under dessa förhållanden. När konvergens inträffar i en viss miljö kan det antas att nämnda egenskap är av typen adaptiv. Ytterligare studier behövs dock för att verifiera funktionens funktion, genom bevis som stöder att det faktiskt ökar kondition av populationen.
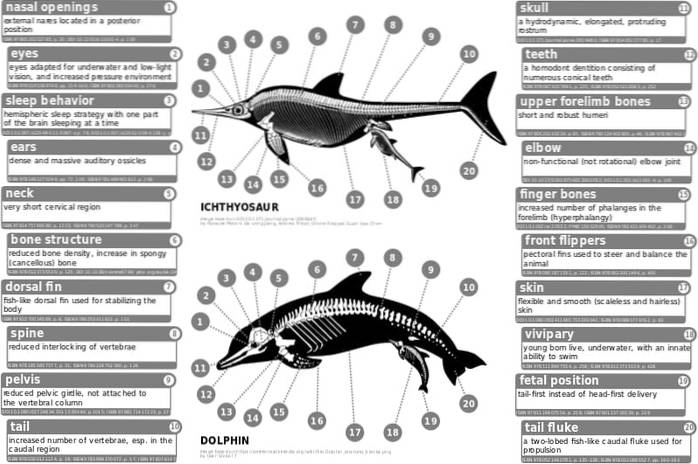
Bland de mest anmärkningsvärda exemplen på konvergent utveckling kan vi nämna flygningen hos ryggradsdjur, ögat hos ryggradsdjur och ryggradslösa djur, spindeln bildar bland annat fisk och vattenlevande däggdjur..
Artikelindex
Låt oss föreställa oss att vi träffar två personer som fysiskt är ganska lika. De har båda samma höjd, ögonfärg och hårfärg. Deras funktioner liknar också. Vi antar antagligen att de två personerna är syskon, kusiner eller kanske avlägsna släktingar..
Trots detta skulle det inte vara en överraskning att få veta att det inte finns någon nära familjär relation mellan människorna i vårt exempel. Detsamma gäller i stor skala i evolutionen: ibland delar liknande former inte en nyare gemensam förfader.
Det vill säga, under hela evolutionen kan egenskaper som liknar i två eller flera grupper förvärvas på olika sätt. Självständig.
Biologer använder två allmänna definitioner för evolutionär konvergens eller konvergens. Båda definitionerna kräver att två eller flera linjer utvecklar tecken som liknar varandra. Definitionen integrerar vanligtvis termen "evolutionär oberoende", även om den är implicit.
Men definitionerna skiljer sig åt i den specifika evolutionära processen eller mekanismen som krävs för att få mönstret..
Några definitioner av konvergens som saknar mekanism är följande: "oberoende utveckling av liknande egenskaper från ett förfäders drag" eller "utveckling av liknande egenskaper i oberoende evolutionära släkter".
Däremot föredrar andra författare att integrera en mekanism i begreppet coevolution för att förklara mönstret.
Till exempel "den oberoende utvecklingen av liknande egenskaper i avlägset besläktade organismer på grund av uppkomsten av anpassningar till liknande miljöer eller livsformer".
Båda definitionerna används ofta i vetenskapliga artiklar och i litteraturen. Den avgörande tanken bakom evolutionär konvergens är att förstå att den gemensamma förfadern till de inblandade linjerna hade ett initialt tillstånd annorlunda.
Efter definitionen av konvergens som inkluderar en mekanism (nämndes i föregående avsnitt) förklarar detta likheten mellan fenotyperna tack vare likheten med de selektiva tryck som taxorna upplever..
Mot bakgrund av evolutionen tolkas detta i termer av anpassningar. Det vill säga de egenskaper som erhålls tack vare konvergens är anpassningar för nämnda miljö, eftersom det på något sätt skulle öka dess kondition.
Det finns dock fall där evolutionskonvergens uppträder och egenskapen inte är anpassningsbar. Det vill säga de berörda linjerna är inte under samma selektiva tryck.
I litteraturen är det vanligt att skilja mellan konvergens och parallellitet. Vissa författare använder det evolutionära avståndet mellan grupperna som ska jämföras för att separera de två begreppen.
Den upprepade utvecklingen av ett drag i två eller flera grupper av organismer anses vara en parallellism om liknande fenotyper utvecklas i besläktade linjer, medan konvergens involverar utvecklingen av liknande egenskaper i separata eller relativt avlägsna linjer..
En annan definition av konvergens och parallellism syftar till att separera dem när det gäller de utvecklingsvägar som är involverade i strukturen. I detta sammanhang producerar konvergent evolution liknande egenskaper genom olika utvecklingsvägar, medan parallell evolution gör det genom liknande vägar..
Skillnaden mellan parallell och konvergent utveckling kan emellertid vara kontroversiell och blir ännu mer komplicerad när vi går ner till identifieringen av den molekylära grunden för det aktuella draget. Trots dessa svårigheter är de evolutionära konsekvenserna relaterade till båda begreppen betydande..
Även om urval gynnar liknande fenotyper i liknande miljöer, är det inte ett fenomen som kan tillämpas i alla fall..
Likheter, ur form och morfologi, kan leda organismer att konkurrera med varandra. Som en konsekvens av detta gynnar urval divergens mellan arter som samexisterar lokalt, vilket skapar en spänning mellan de grader av konvergens och divergens som förväntas för en viss livsmiljö..
Individer som är nära och har betydande nischöverlappning är de mest kraftfulla konkurrenterna - baserat på deras fenotypiska likhet, vilket får dem att utnyttja resurser på liknande sätt.
I dessa fall kan avvikande urval leda till ett fenomen som kallas adaptiv strålning, där en härstamning ger upphov till olika arter med en stor mångfald av ekologiska roller på kort tid. Förhållandena som främjar adaptiv strålning inkluderar bland annat miljömässig heterogenitet, frånvaro av rovdjur..
Adaptiv strålning och konvergent utveckling betraktas som två sidor av samma "evolutionära mynt".
För att förstå skillnaden mellan evolutionär konvergens och paralleller uppstår en mycket intressant fråga: när naturligt urval gynnar utvecklingen av liknande egenskaper, förekommer det under samma gener eller kan det involvera olika gener och mutationer som resulterar i liknande fenotyper?
Baserat på de bevis som genererats hittills verkar svaret på båda frågorna vara ja. Det finns studier som stöder båda argumenten.
Även om det hittills inte finns något konkret svar på varför vissa gener ”återanvänds” i evolutionär evolution, finns det empiriska bevis som syftar till att belysa saken..
Till exempel har den upprepade utvecklingen av blomningstider i växter, insekticidresistens hos insekter och pigmentering hos ryggradsdjur och ryggradslösa djur visat sig inträffa genom förändringar som involverar samma gener..
Men för vissa egenskaper kan endast ett litet antal gener förändra egenskaperna. Ta synen: förändringar i färgvision måste nödvändigtvis inträffa i förändringar relaterade till opsin-gener.
I andra egenskaper är generna som kontrollerar dem däremot fler. Cirka 80 gener är involverade i växternas blomningstider, men förändringar har bara visats under hela utvecklingen hos några få.
1997 undrade Moore och Willmer hur vanligt fenomenet konvergens är.
För dessa författare är denna fråga obesvarad. De hävdar att det, baserat på de hittills beskrivna exemplen, finns relativt höga nivåer av konvergens. De hävdar dock att det fortfarande finns en betydande underskattning av evolutionär konvergens i organiska varelser.
I evolutionböcker hittar vi ett dussin klassiska exempel på konvergens. Om läsaren vill utöka sin kunskap om ämnet kan han läsa McGhees bok (2011), där han hittar många exempel i olika grupper av livets träd.
I organiska varelser är ett av de mest häpnadsväckande exemplen på evolutionär konvergens uppkomsten av flygning i tre linjer av ryggradsdjur: fåglar, fladdermöss och de nu utdöda pterodaktylerna..
I själva verket går konvergens i nuvarande flygande ryggradsdjurgrupper utöver att ha ändat frambenen till strukturer som tillåter flygning..
En serie fysiologiska och anatomiska anpassningar delas mellan båda grupperna, såsom det kännetecknande att ha kortare tarmar, som antas, minskar individens massa under flygningen, vilket gör det billigare och mer affektivt..
Ännu mer överraskande har olika forskare funnit evolutionära konvergenser inom fladdermus- och fågelgrupper på familjens nivå..
Fladdermöss i familjen Molossidae liknar till exempel familjemedlemmarna Hirundinidae (svalor och allierade) hos fåglar. Båda grupperna kännetecknas av snabb flygning, i höga höjder, med liknande vingar..
På liknande sätt konvergerar medlemmar av familjen Nycteridae i olika avseenden med passerine fåglar (Passeriformes). Båda flyger i låga hastigheter och har förmågan att manövrera genom vegetationen.
Ett enastående exempel på evolutionär konvergens finns när man analyserar två grupper av däggdjur: aye-igår och ekorrarna..
Idag är aye-aye (Daubentonia madagascariensis) klassificeras som en lemuriform primat endemisk till Madagaskar. Deras ovanliga kost består i grunden av insekter.
Således har aye-aye anpassningar som har varit relaterade till dess trofiska vanor, såsom akut hörsel, förlängning i långfingret och tänder med växande framtänder..
När det gäller protesen liknar den i flera avseenden hos en gnagare. Inte bara i snittets utseende, de delar också en utomordentligt liknande tandformel.
Utseendet mellan de två taxorna är så slående att de första taxonomerna klassificerade aye-aye, tillsammans med de andra ekorrarna, i släktet Sciurus.



Ingen har kommenterat den här artikeln än.