De globosider De är en typ av sfingolipider som tillhör den heterogena familjen glykosfingolipider och kännetecknas av att i sina strukturer har en polär grupp bestående av komplexa strukturerade glykaner kopplade till ett ceramidskelett genom en B-glykosidbindning.
De klassificeras inom "glob" -serien av glykosfingolipider genom närvaron av en central struktur av den allmänna formen Galα4Galβ4GlcβCer, och deras nomenklatur baseras i allmänhet på antalet och typen av sockerrester av polhuvudena.
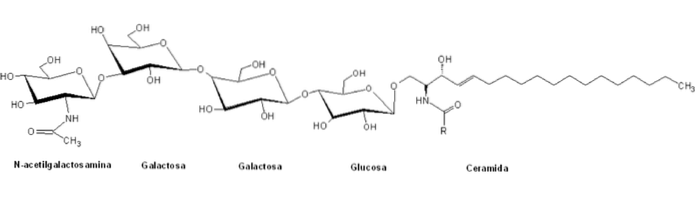
Till skillnad från andra sfingolipider är globosider normala beståndsdelar i cellmembran i icke-nervsystem i många däggdjur. Till exempel njurarna, tarmarna, lungorna, binjurarna och röda blodkroppar.
Liksom alla membranlipider har globosider viktiga strukturfunktioner vid bildning och ordning av lipid dubbelskikt..
Men till skillnad från deras sura eller fosforylerade motsvarigheter är globosidernas funktion inte så relaterad till produktionen av signalmolekyler, utan snarare till deras deltagande som en del av glykokonjugat i plasmamembranet..
Artikelindex
De delar vissa strukturella och funktionella likheter med de andra medlemmarna i gruppen glukosfingolipider: cerebrosider, gangliosider och sulfatider; inklusive sammansättningen av huvudskelettet och biprodukterna av dess metabolism.
Globosider skiljer sig emellertid från sura glykosfingolipider (såsom gangliosider) angående laddningen av deras kolhydratpolargrupper, eftersom de är elektriskt neutrala vid fysiologiskt pH, vilket verkar ha starka konsekvenser för deras funktioner som en del av den extracellulära matrisen..
Dessa polära huvudgrupper har normalt mer än två sockermolekyler, varav vanligtvis D-glukos, D-galaktos och N-acetyl-D-galaktosamin, och i mindre utsträckning fukos och N-acetylglukosamin.
Som med andra sfingolipider kan globosider vara mycket olika molekyler, antingen med hänsyn till de multipla kombinationer av fettsyror som är bundna till sfingosinskelettet eller de möjliga variationerna av oligosackaridkedjorna i den hydrofila delen..
Vägen börjar med syntesen av ceramid i endoplasmatisk retikulum (ER). Sfingosinskelettet bildas först genom kondensation av en L-serin och en palmitoyl-CoA.
Ceramid genereras därefter genom verkan av ceramidsyntasenzymer, som kondenserar en annan molekyl fettsyra-CoA med ryggraden i sfingosin vid kolet vid position 2.
Fortfarande i ER kan de producerade ceramiderna modifieras genom tillsats av en galaktosrest för att bilda galacto ceramides (GalCer), eller de kan istället transporteras till Golgi-komplexet antingen genom inverkan av ceramidöverföringsproteiner (CERT) eller medelst av vesikulär transport.
I Golgi-komplexet kan ceramider glykosyleras för att producera glukoceramider (GlcCer).
GlcCer produceras på den tidiga Golgis cytosoliska ansikte. Det kan sedan transporteras till komplexets luminala yta och därefter glykosyleras av specifika glykosidasenzymer som genererar mer komplexa glykosfingolipider..
De vanliga föregångarna till alla glykosfingolipider syntetiseras i Golgi-komplexet genom inverkan av glykosyltransferaser från GalCer eller GlcCer.
Dessa enzymer överför specifika kolhydrater från lämpliga nukleotidsocker: UDP-glukos, UDP-galaktos, CMP-sialinsyra, etc..
När GlcCer passerar genom Golgi vesikulärt människosmugglingssystem galaktosyleras det för att producera laktosylceramid (LacCer). LacCer är grenpunkten från vilken föregångarna till de andra glykosfingolipiderna syntetiseras, det vill säga molekylen till vilken mer neutrala polära sockerrester sedan tillsätts. Dessa reaktioner katalyseras av specifika globosidsyntaser.
Dessa lipider finns främst i mänskliga vävnader. Liksom många glykosfingolipider berikas globosider på utsidan av plasmamembranet i många celler..
De är särskilt viktiga i humana erytrocyter, där de representerar den huvudsakliga typen av glykolipid på cellytan..
Dessutom, som nämnts ovan, är de en del av uppsättningen glykokonjugat i plasmamembranen i många icke-nervorgan, huvudsakligen njurarna..
Globosidernas funktioner har hittills inte blivit helt klarlagda, men det är känt att vissa arter ökar cellproliferation och rörlighet, i motsats till hämningen av dessa händelser orsakade av vissa gangliosider..
En tetra-glykosylerad globosid, Gb4 (GalNAcβ3Galα4Galβ4GlcβCer), fungerar i plats-känslig igenkänning av strukturella störningar av erytrocyter under celladhesionsprocesser..
Nya studier har bestämt involveringen av Gb4 i aktiveringen av ERK-proteiner i karcinomcellinjer, vilket kan betyda dess deltagande i tumörinitiering. Dessa proteiner tillhör det mitogenaktiverade proteinkinas (MAPK) signalkaskad, bestående av elementen Raf, MEK och ERK..
Deras deltagande som receptorer för vissa bakterietoxiner i Shiga-familjen har rapporterats, specifikt globosiden Gb3 (Galα4Galβ4GlcβCer), även känd som CD77, uttryckt i omogna B-celler; också som receptorer för HIV-vidhäftningsfaktor (gp120) och verkar ha konsekvenser för vissa typer av cancer och andra sjukdomar.
Det finns många typer av lipidos hos människor. Globosider och deras metaboliska vägar är särskilt relaterade till två sjukdomar: Fabrys sjukdom och Sandhoffs sjukdom..
Det hänvisar till en könsbunden ärftlig systemisk störning, först sett hos patienter med flera lila fläckar i navelregionen. Det påverkar organ som njurar, hjärta, ögon, extremiteter, en del av mag-tarmkanalen och nervsystemet.
Det är produkten av en metabolisk defekt i enzymet ceramid trihexosidas, som ansvarar för hydrolys av trihexosiceramid, en mellanprodukt i katabolismen av globosider och gangliosider, vilket orsakar en ansamling av dessa glykolipider i vävnaderna.
Denna patologi beskrivs ursprungligen som en variant av Tay-Sachs sjukdom, relaterad till metabolismen av gangliosider, men detta presenterar också ackumuleringen av globosider i inälvorna. Det är en ärftlig sjukdom med autosomala recessiva mönster som gradvis förstör nervceller och ryggmärg.
Det har att göra med frånvaron av formerna A och B av enzymet β-N-acetylhexosaminidas på grund av mutationer i genen HEXB. Dessa enzymer är ansvariga för ett av nedbrytningsstegen hos vissa glykosfingolipider.



Ingen har kommenterat den här artikeln än.