De profas det är det första steget av celldelning genom mitos och meios. Det är fasen efter DNA-syntesstadiet (S-fasen i cellcykeln). I detta skede når kromosomerna en hög grad av kondens och individualitet.
I meios finns det två proteaser, som skiljer sig mycket från varandra och från mitos. Endast i meiotisk profas I, till exempel, sker rekombination. Denna fas är uppdelad i olika steg: leptoten, zygoten, pachyten, diploten och diakinesis..

Under profas, utöver kondensationen som uppnås av de duplicerade kromosomerna, sker förändringsprocesser på den extra kärnkraftsnivån. Den viktigaste cytoplasmiska händelsen under profas är bildandet av den akromatiska spindeln vid varje cellpol. Detta gör att kromosomerna kan mobiliseras i successiva skeden av celldelning för att säkerställa korrekt segregering..
Det finns viktiga skillnader mellan celldelningar i djurceller och växtceller. Vissa kommer att nämnas senare. Sammantaget sker emellertid en fullständig omorganisation av cellen.
Därför fokuserar mitos och meios på ödet för DNA och kärnan. Men sanningen är att när en cell delar upp delar den allt och allt deltar i processen.
Således genomgår alla cellulära komponenter radikala förändringar under mitos och meios. Det endoplasmiska nätverket och Golgi-komplexet verkar försvinna: de ändrar dock bara sin struktur. Mitokondrier och kloroplaster delar sig också, vilket ger upphov till nya organeller..
Artikelindex

Djurceller har en enda centriole. Efter avslutad DNA-syntes som förberedelse för mitos förbereder centriolen sig också för att dela sig.
Centrioler består av ett par identiska strukturer som kallas diplosomer, vinkelräta mot varandra. Dessa separerar, och var och en kommer att vara en form för uppkomsten av en ny. Syntesen av den nya diplosomen sker när varje gammal diplosom migrerar till motsatta poler i cellen..
Den andra definierande händelsen av profas, och en som delas med växtceller, är kromatinkomprimering. Detta är kanske det mest anmärkningsvärda cytologiska elementet i profas under celldelning..
DNA når en hög grad av komprimering och för första gången observeras det som en morfologiskt individualiserad kromosom.
De komprimerade kromosomerna involverar systerkromatiderna för var och en av dem, fortfarande förenade av samma centromer. Även om den här centromeren verkligen är dubbel, beter den sig som en enkel.
Kromosomerna kommer att ses som ett X, eftersom de är två kopieringskromatider fästa i samma centrum. Därför kommer varje cell i profas att ha ett dubbelt antal kromatider, jämfört med ett antal centromerer som är lika med antalet '2n' av arten..
Det vill säga, en profos mitotisk cell är diploid efter antal centromerer, men tetraploid (4n) efter antal kromatider..
I växtceller finns det en fas före profas som kallas preprofas. Som förberedelse för celldelning sönderfaller den stora cellvakuolen.
Tack vare detta bildas ett fritt eller ledigt cytoplasmatiskt band, kallat fragmosom. Detta gör att växtcellkärnan kan positionera sig mot cellens ekvatorn.
Dessutom kollapsar den kortikala organisationen av mikrotubuli mot samma plats. Detta kommer att ge upphov till det som kallas pre-prophase band (BPP)..

Prefasbandet kommer att visas först som en ring, men slutar täcka kärnan. Det vill säga mikrotubuli som leder cellmembranet internt kommer alla att mobilisera mot fragmosomen.
Därefter kommer preprofasbandet som omger ekvatorkärnan att tillåta att lokalt organisera den plats där fragmoplasten som kommer att ersätta den så småningom kommer att visas..
Dynamiskt sett kommer växtcellens mikrotubuli att passera från en fas till en annan utan uppenbara övergångar. Det vill säga från kortikalt arrangemang till fragmosom och därifrån till fragmoplast.
Platsen för alla dessa strukturella förändringar i växtcellen är densamma där avlagringen av cellplattan kommer att ske. Och därför representerar det planet där cellen kommer att dela sig.
För allt annat är växtprofasen identiskt med det som observeras i djurcellernas profas.
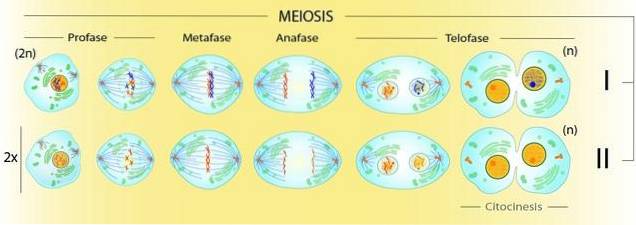
Endast i profas I om meios uppstår genetisk rekombination. Därför kräver bildandet av komplexa strukturer mellan kromosomer att det finns två indelningar i meios.
Med tidigare DNA-syntes framställdes systerkromatiderna på varje kromosom. Med deras komprimering har vi dubbla kromosomer som i meios dessutom parar mellan homologer.
Detta leder till generering av bivalenter (två interagerande homologa kromosomer). Eftersom var och en dupliceras talar vi faktiskt om tetrader. Det vill säga av kromatidtetrader förenade i en struktur som måste lösas med hjälp av två celldelningar.
I den första kommer de homologa kromosomerna att separeras, medan i den andra måste systerkromatiderna separeras.
I meiotisk profas I är systerkromatider organiserade på kompakta proteinhaltiga strukturer som utgör den centrala kromosomala axeln..
På denna axel kommer det synaptonemiska komplexet (CS) att bildas, vilket kommer att hålla de homologa kromosomerna i parning. Under profas I tillåter det synaptonemiska komplexet att homologa kromosomer kan komma in i synapser..
I dessa steg kan korsningspunkter bildas, synliga som chiasmas, där den genetiska rekombinationsprocessen kommer att äga rum. Det vill säga det fysiska utbytet mellan de deltagande DNA-molekylerna som definierar pachytenen.
Profas II föregås inte av tidigare DNA-syntes. Här ärvdes dubbla kromosomer förenade med samma centromer (dubbel). Detta beror på att DNA-syntes, både i mitos och meios, endast sker i S (syntes) -fasen i cellcykeln..
I denna andra division kommer vi att ha fyra meiocyter. En meiocyt är en cell som är en produkt av en meiotisk uppdelning.
Profas II kommer därför att ha ansvaret för separationen av systerkromatider från kromosomer som ärvs från profas I. Därför kommer varje meiocyt i slutet av den meiotiska processen att ha den haploida uppsättningen kromosomer av arten.



Ingen har kommenterat den här artikeln än.