
De pachyten eller paquinema är den tredje etappen av meiotisk profas I; i den verifieras rekombinationsprocessen. I mitos finns ett profas och i meios två: profas I och profas II.
Tidigare, med undantag av profas II, duplicerades kromosomerna, var och en gav upphov till en systerkromatid. Men bara i profas parar jag ihop homologer (dubbletter) och bildar bivalenter.

Termen paquiteno kommer från grekiska och betyder "tjocka trådar". Dessa "tjocka trådar" är de ihopkopplade homologa kromosomerna som efter duplicering bildar tetrader. Det vill säga fyra "trådar" eller strängar som gör att varje kromosom ser tjockare ut.
Det finns unika aspekter av meiotisk profas I som förklarar de inneboende egenskaperna hos pachyten. Endast i meiosprofas I pachyten rekombineras kromosomer.
För att göra detta verifieras erkännandet och matchningen av homologerna. Liksom vid mitos måste kromatiderna dupliceras. Men det är bara i meiosis I pachyten som bandbyteskomplex bildas, som vi kallar chiasmata..
I dem förekommer det som definierar den rekombinanta kraften hos meios: korsningen mellan kromatider av homologa kromosomer.
Hela processen för DNA-utbyte är möjlig tack vare det tidigare synaptonemiska komplexet. Detta multiproteinkomplex gör att homologa kromosomer kan paras (synaps) och rekombineras..
Artikelindex
Det synaptonemiska komplexet (CS) är proteinramen som möjliggör änd-till-slut-korsning mellan homologa kromosomer. Det förekommer endast under pachytenen av meios I och är den fysiska grunden för kromosomparning. Med andra ord är det det som gör att kromosomer kan synapsas och rekombineras..
Det synaptonemiska komplexet är mycket bevarat bland eukaryoter som genomgår meios. Därför är det evolutionärt mycket gammalt, och strukturellt och funktionellt ekvivalent i alla levande saker..
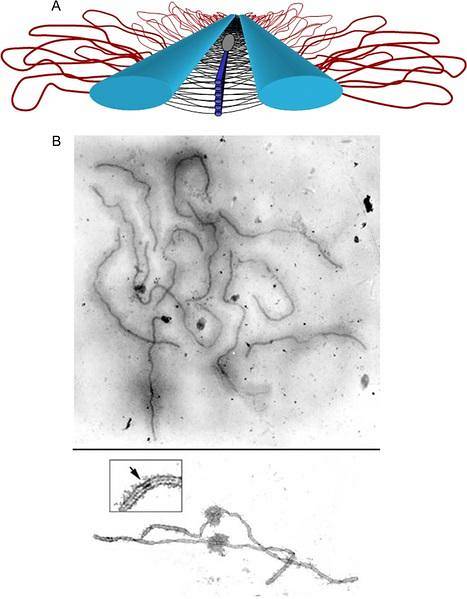
Den består av ett centralt axiellt element och två laterala element som upprepas som tänderna på en dragkedja eller förslutning..
Det synaptonemiska komplexet bildas från specifika punkter på kromosomerna under zygoten. Dessa platser är kollinära med de där DNA-brott inträffar där synapser och rekombination kommer att upplevas i pachytenen..
Under pachytenen har vi därför en sluten dragkedja. I denna konformation definieras specifika punkter där DNA-band kommer att utbytas i slutet av scenen..
Det meiotiska synaptonemiska komplexet innehåller många strukturella proteiner som också finns under mitos. Dessa inkluderar topoisomeras II, kondensiner, kohesiner, såväl som kohesinassocierade proteiner..
Utöver dessa finns också proteiner som är specifika och unika för meios, tillsammans med proteiner från det rekombinanta komplexet..
Dessa proteiner är en del av rekombinosomen. Denna struktur grupperar alla proteiner som krävs för rekombination. Uppenbarligen bildas rekombinosomen inte på delningsställena utan rekryteras, redan bildade, mot dem.
Chiasms är de synliga morfologiska strukturerna på kromosomer där korsningar förekommer. Med andra ord, den fysiska manifestationen av utbytet av DNA-band mellan två homologa kromosomer. Chiasms är de särskiljande cytomorfologiska märkena för pachyten.
I all meios måste minst en chiasm per kromosom förekomma. Detta innebär att varje gamet är rekombinant. Tack vare detta fenomen kunde de första genetiska kartorna baserade på koppling och rekombination dras och föreslås..
Å andra sidan orsakar bristen på chiasmer och därför crossover, snedvridningar på nivån av kromosomal segregering. Rekombination under pachyten fungerar sedan som en kvalitetskontroll av meiotisk segregering..
Men evolutionärt sett genomgår inte alla organismer rekombination (till exempel manliga fruktflugor). I dessa fall fungerar andra mekanismer för kromosomavskiljning som inte är beroende av rekombination..
Vid utträde av zygoten är det synaptonemiska komplexet helt bildat. Detta kompletteras med genereringen av dubbelband-DNA-avbrott från vilka delningsfilter verifieras..
Dubbel DNA-brott tvingar cellen att reparera dem. I processen för DNA-reparation rekryterar cellen rekombinosomen. Bandbyte används och som ett resultat erhålls rekombinanta celler.
När det synaptonemiska komplexet är helt bildat sägs pachytenen börja.
Bivalenterna i synapser i pachytenen interagerar i princip genom det axiala elementet i det synaptonemiska komplexet. Varje kromatid är organiserad i en looporganisation, vars bas är det centrala axiella elementet i det synaptonemiska komplexet..
Det axiella elementet hos varje motpart kommer i kontakt med det andra genom sidoelementen. Systerkromatidaxlarna är mycket komprimerade och deras kromatinslingor dyker upp från det centrala axiella elementet. Ögonavstånd (~ 20 per mikrometer) bevaras evolutionärt över alla arter.
Mot slutet av pachytenen är korsningar uppenbara från några av dubbelbands-DNA-brytplatserna. Utseendet på delarna signalerar också början på det synaptonemiska komplexet..
Homologa kromosomer blir mer kondenserade (ser mer individuella ut) och börjar separeras, förutom i chiasmata. När detta händer slutar pachytenen och diploten börjar..
Föreningen mellan rekombinosomen och axlarna i det synaptonemiska komplexet kvarstår under hela synapsen. Särskilt i rekombinogena korsningar till slutet av pachytenen, eller något bortom.


Ingen har kommenterat den här artikeln än.