De ribos Det är ett socker med fem kol som finns i ribonukleosider, ribonukleotider och deras derivat. Det kan hittas med andra namn såsom β-D-ribofuranos, D-ribos och L-ribos.
Nukleotider är "byggstenarna" i ribonukleinsyran (RNA). Varje nukleotid består av en bas som kan vara adenin, guanin, cytosin eller uracil, en fosfatgrupp och ett socker, ribos.

Denna typ av socker är särskilt riklig i muskelvävnader, där det är associerat med ribonukleotider, särskilt med adenosintrifosfat eller ATP, vilket är viktigt för muskelfunktion..
D-ribos upptäcktes 1891 av Emil Fischer, och sedan dess har mycket uppmärksamhet ägnats åt dess fysikalisk-kemiska egenskaper och dess roll i cellulär metabolism, det vill säga som en del av skelettet av ribonukleinsyra, ATP och olika koenzymer..
Först erhölls detta endast från hydrolys av jäst-RNA, tills det på 1950-talet lyckades syntetiseras från D-glukos i mer eller mindre tillgängliga kvantiteter, vilket möjliggjorde industrialisering av dess produktion..
Artikelindex
Ribos är en aldopentos som ofta extraheras som en ren kemisk förening i form av D-ribos. Det är en organisk substans löslig i vatten med ett vitt och kristallint utseende. Att vara kolhydrat har ribos polära och hydrofila egenskaper.
Ribos uppfyller den vanliga kolhydratregeln: den har samma antal kol- och syreatomer och två gånger så många i väteatomer..
Genom kolatomerna i positionerna 3 eller 5 kan detta socker bindas till en fosfatgrupp, och om det binder till en av kvävebaserna i RNA bildas en nukleotid.
Det vanligaste sättet att hitta ribos i naturen är som D-ribos och 2-deoxi-D-ribos, dessa är komponenter i nukleotider och nukleinsyror. D-ribos är en del av ribonukleinsyra (RNA) och 2-deoxi-D-ribos av deoxiribonukleinsyra (DNA).
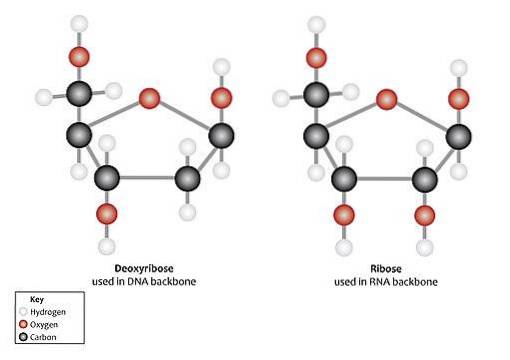
Vid nukleotider är båda typerna av pentos i β-furanosform (sluten femkantig ring).
I lösning är fri ribos i jämvikt mellan aldehydformen (öppen kedja) och den cykliska β-furanosformen. Emellertid innehåller RNA endast den cykliska formen β-D-ribofuranos. Den biologiskt aktiva formen är vanligtvis D-ribos.
Ribos är ett socker som härrör från glukos som tillhör gruppen aldopentoser. Dess molekylformel är C5H10O5 och har en molekylvikt på 150,13 g / mol. Eftersom det är ett monosackaridsocker, separerar dess hydrolys molekylen i dess funktionella grupper.
Den har, som dess formel indikerar, fem kolatomer som kan återfinnas cykliskt som en del av fem- eller sexledade ringar. Detta socker har en aldehydgrupp vid kol 1 och en hydroxylgrupp (-OH) vid kolatomer från position 2 till position 5 i pentosringen.
Ribosmolekylen kan representeras i Fisher-projektionen på två sätt: D-ribos eller L-ribos, med form L som stereoisomer och enantiomer av form D och vice versa..
Klassificeringen av D- eller L-formen beror på orienteringen av hydroxylgrupperna i den första kolatomen efter aldehydgruppen. Om denna grupp är orienterad mot höger sida motsvarar molekylen i Fishers representation D-ribos, annars om den är mot vänster sida (L-ribos).
Haworth-projektionen av ribos kan representeras i två ytterligare strukturer beroende på orienteringen av hydroxylgruppen på kolatomen som är anomer. I β-positionen är hydroxylen orienterad mot molekylens topp, medan α-positionen orienterar hydroxylen mot botten.

Enligt Haworth-projektionen kan det således finnas fyra möjliga former: β-D-ribos, α-D-ribos, β-L-ribos eller α-L-ribos.
När fosfatgrupper är bundna till ribos, kallas dessa ofta som α, β och Ƴ. Hydrolys av nukleosidtrifosfat ger den kemiska energin för att driva en mängd olika cellulära reaktioner.
Det har föreslagits att ribosfosfat, produkt från nedbrytning av ribonukleotider, är en av de viktigaste föregångarna till furan och tiofenoler, som är ansvariga för den karakteristiska lukten av kött..
Ribosens kemiska plasticitet gör att molekylen är involverad i den stora majoriteten av biokemiska processer inuti cellen, vissa som översättning av DNA, syntes av aminosyror och nukleotider etc..
Ribos fungerar ständigt som en kemisk vehikel inuti cellen, eftersom nukleotider kan ha en, två eller tre fosfatgrupper kovalent kopplade till varandra genom vattenfria bindningar. Dessa är kända som nukleosiderna mono-, di- och trifosfat..
Bindningen mellan ribos och fosfat är av estertyp, hydrolysen av denna bindning frigör ungefär 14 kJ / mol under standardbetingelser, medan den för var och en av anhydridbindningarna frigör cirka 30 kJ / mol.
I ribosomer kan till exempel 2'-hydroxylgruppen i ribos bilda en vätebindning med de olika aminosyrorna, en sammanslutning som möjliggör syntes av proteiner från tRNA i alla kända levande organismer.
Giftet för de flesta ormar innehåller ett fosfodiesteras som hydrolyserar nukleotider från 3'-änden som har en fri hydroxyl, vilket bryter bindningarna mellan 3'-hydroxylen av ribos eller deoxiribos.
I medicinska sammanhang används det för att förbättra prestanda och träningskapacitet genom att öka muskelenergin. Kroniskt trötthetssyndrom behandlas också med denna sackarid, liksom fibromyalgi och vissa kranskärlssjukdomar..
I förebyggande termer används den för att förhindra muskeltrötthet, kramper, smärta och stelhet efter träning hos patienter med ärftlig störning av myoadenylatdeaminasbrist eller AMP-deaminasbrist..



Ingen har kommenterat den här artikeln än.