
De Golgiapparat, Även känt som Golgi-komplex, det är en organell cell i membran som bildas av en uppsättning plana blåsor staplade ihop; Dessa säckar har vätska inuti. Finns i en mängd olika eukaryoter, inklusive djur, växter och svampar.
Denna organell ansvarar för bearbetning, förpackning, klassificering, distribution och modifiering av proteiner. Dessutom har den en roll i syntesen av lipider och kolhydrater. Å andra sidan sker i växter syntesen av komponenterna i cellväggen i Golgi-apparaten..
Golgi-apparaten upptäcktes 1888 medan nervceller studerades; dess upptäckare, Camillo Golgi, vann Nobelpriset. Strukturen kan detekteras genom silverkromatfärgning.
Först var orgelns existens tveksamt för tidens vetenskapsmän och de tillskrev Golgi-observationerna till enkla artefakter, produkter med de tekniker som används.
Artikelindex
Golgi-apparaten är en eukaryot organell av membranös natur. Det liknar påsar i staplar, även om organisationen kan variera beroende på celltyp och organismen. Det ansvarar för modifiering av proteiner efter översättning.
Till exempel kan en del kolhydrater tillsättas för att bilda ett glykoprotein. Denna produkt förpackas och distribueras till cellfacket där det behövs, såsom membran, lysosomer eller vakuoler; det kan också skickas utanför cellen. Det deltar också i syntesen av biomolekyler.
Cytoskelettet (specifikt aktin) bestämmer dess läge, och i allmänhet är komplexet beläget i ett område av det inre av cellen nära kärnan och centrosomen.
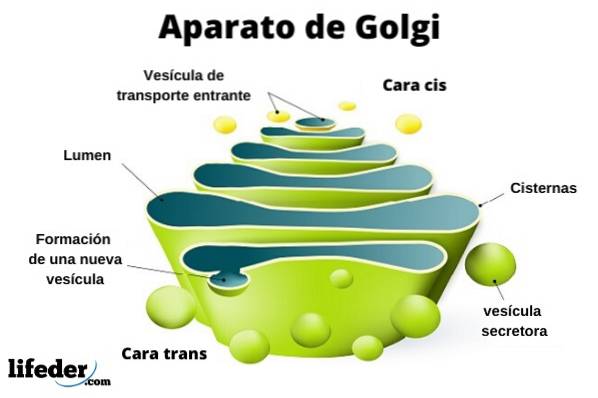
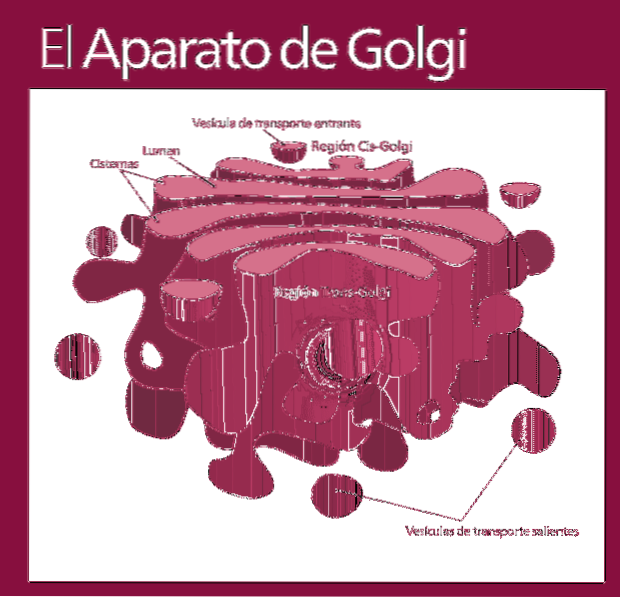
Golgi-komplexet består av en uppsättning platta, fenestrerade, skivformade säckar, kallade Golgian-cisterner, med varierande tjocklek.
Dessa säckar staplas i grupper om fyra eller sex cisterner. I en däggdjurscell kan du hitta mellan 40 och 100 batterier anslutna till varandra.
Golgi-komplexet presenterar en intressant egenskap: det finns polaritet när det gäller struktur och funktion.
Du kan skilja mellan cis-ansiktet och trans-ansiktet. Den första är relaterad till inträde av proteiner och ligger nära det endoplasmatiska retikulumet. Den andra är utgångsytan eller produktsekretionen; De bildas av en eller två tankar som har en rörform.
Tillsammans med denna struktur finns blåsor som utgör transportsystemet. Högarna med säckar är fästa vid varandra i en struktur som påminner om en båge eller datum.
Hos däggdjur fragmenteras Golgi-komplexet i flera vesiklar under celldelningsprocesser. Blåsorna passerar till dottercellerna och tar igen den traditionella formen av komplexet.
Organisationen av komplexet är inte vanligt i alla grupper av organismer. I vissa celltyper är komplexet inte strukturerat som uppsättningar cisterner staplade i grupper; tvärtom är de placerade individuellt. Ett exempel på denna organisation är svampens Saccharomyces cerevisiae.
I vissa encelliga organismer, såsom toxoplasma eller trypanosom, närvaron av endast en membranhög har rapporterats.
Alla dessa undantag tyder på att staplingen av strukturerna inte är nödvändig för att de ska kunna utföra deras funktion, även om närheten mellan påsarna gör transportprocessen mycket effektivare..
På samma sätt saknar vissa basala eukaryoter dessa cisternaer; till exempel svamp. Detta bevis stöder teorin att apparaten uppträdde i en senare släktlinje än de första eukaryoterna..
Funktionellt är Golgi-komplexet uppdelat i följande fack: cis-nätverket, de staplade säckarna - som i sin tur är uppdelade i mitt- och trans-underavdelningen - och trans.
Molekylerna som kommer att modifieras går in i Golgi-komplexet i samma ordning (cis-nätverk följt av underkomponenter som slutligen utsöndras i trans-nätverket).
De flesta reaktioner förekommer i de mest aktiva zonerna: trans- och mittdelarna.
Golgi-komplexet har som huvudfunktion post-translationell modifiering av proteiner tack vare de enzymer de har inuti.
Dessa modifieringar inkluderar processer för glykosylering (tillsats av kolhydrater), fosforylering (tillsats av en fosfatgrupp), sulfatering (tillsats av en fosfatgrupp) och proteolys (nedbrytning av proteiner).
Dessutom är Golgi-komplexet involverat i syntesen av specifika biomolekyler. Var och en av dess funktioner beskrivs i detalj nedan:
I Golgi-apparaten sker modifiering av ett protein till ett glykoprotein. Det typiska sura pH i det inre av organellen är avgörande för att denna process ska ske normalt..
Det är ett konstant utbyte av material mellan Golgi-apparaten med det endoplasmiska retikulumet och lysosomerna. I det endoplasmiska retikulumet genomgår också proteiner modifieringar; dessa inkluderar tillsatsen av en oligosackarid.
När dessa molekyler (N-oligosackarider) kommer in i Golgi-komplexet får de en rad ytterligare modifieringar. Om denna molekyls öde ska bäras utanför cellen eller tas emot i plasmamembranet inträffar speciella modifieringar.
Dessa modifieringar inkluderar följande steg: avlägsnande av tre mannosrester, tillsats av N-acetylglukosamin, avlägsnande av två mannoser och tillsats av fukos, ytterligare två N-acetylglukosamin-, tre galaktos- och tre sialinsyrarester..
Däremot modifieras proteinerna som är avsedda för lysosomer på följande sätt: det finns inget avlägsnande av mannoser som ett första steg; istället sker fosforylering av dessa rester. Detta steg inträffar i cis-regionen i komplexet.
Därefter avlägsnas N-acetylglukosamin-grupperna och lämnar mannoserna med fosfatet tillsatt till oligosackariden. Dessa fosfater indikerar att proteinet måste specifikt riktas mot lysosomer..
Receptorerna som är ansvariga för att känna igen fosfaterna som indikerar deras intracellulära öde finns i transnätverket.
I Golgi-komplexet sker syntesen av glykolipider och sfingomyelin med användning av ceramid (tidigare syntetiserat i det endoplasmatiska retikulum) som ursprungsmolekyl. Denna process strider mot den för resten av fosfolipiderna som utgör plasmamembranet, som härrör från glycerol..
Sfingomyelin är en klass av sfingolipid. Det är en riklig komponent av däggdjursmembran, särskilt nervceller, där de är en del av myelinhöljet.
Efter syntesen transporteras de till sin slutliga plats: plasmamembranet. Deras polära huvuden är placerade mot utsidan av cellytan; dessa element har en specifik roll i celligenkänningsprocesser.
I växtceller bidrar Golgi-apparaten till syntesen av polysackariderna som utgör cellväggen, särskilt hemicellulosa och pektiner. Genom vesikulär transport transporteras dessa polymerer utanför cellen.
I grönsaker är detta steg avgörande och ungefär 80% av aktiviteten i retikulum tilldelas syntesen av polysackarider. Faktum är att hundratals av dessa organeller har rapporterats i växtceller..
De olika biomolekylerna - proteiner, kolhydrater och lipider - överförs till deras cellulära destinationer av Golgi-komplexet. Proteiner har en slags "kod" som är ansvarig för att informera den destination till vilken den tillhör.
De transporteras i vesiklar som lämnar transnätet och färdas till det specifika cellfacket.
Proteiner kan transporteras till membranet genom en specifik konstitutiv väg. Det är därför det finns en kontinuerlig inkorporering av proteiner och lipider i plasmamembranet. Proteiner vars slutdestination är Golgi-komplexet behålls av detta.
Förutom den konstitutiva vägen är andra proteiner avsedda för cellens utsida och uppstår genom signaler från omgivningen, vare sig det är hormoner, enzymer eller neurotransmittorer..
Till exempel, i bukspottkörtelceller, är matsmältningsenzymer förpackade i blåsor som endast utsöndras när närvaron av mat detekteras..
Ny forskning rapporterar förekomsten av alternativa vägar för membranproteiner som inte passerar genom Golgi-apparaten. Men dessa vägar gå förbi "Okonventionellt" debatteras i litteraturen.
Det finns fem modeller för att förklara proteinhandel i apparaten. Det första handlar om trafik av material mellan stabila avdelningar, var och en har nödvändiga enzymer för att uppfylla specifika funktioner. Den andra modellen involverar mognad av cisternerna successivt.
Den tredje föreslår också mognad av påsarna men med införandet av en ny komponent: den rörformiga transporten. Enligt modellen är tubuli viktiga i trafiken i båda riktningarna.
Den fjärde modellen föreslår att komplexet fungerar som en enhet. Den femte och sista modellen är den senaste och hävdar att komplexet är uppdelat i olika fack.
I vissa celltyper har Golgi-komplexet specifika funktioner. Bukspottkörtelns celler har specialiserade strukturer för utsöndring av insulin.
De olika blodtyperna hos människor är ett exempel på olika glykosyleringsmönster. Detta fenomen förklaras av närvaron av olika alleler som kodar för glukotransferas.



Ingen har kommenterat den här artikeln än.