De axoneme Det är en inre cytoskeletal struktur av cilia och flagella baserat på mikrotubuli och som ger dem rörelse. Dess struktur består av ett plasmamembran som omger ett par centrala mikrotubuli och nio par perifera mikrotubuli..
Axonemet ligger utanför cellen och är förankrat inuti cellen med hjälp av baskroppen. Den är 0,2 μm i diameter och dess längd kan variera från 5-10 μm i cilia till flera mm i flagellum hos vissa arter, även om dessa i allmänhet mäter 50-150 μm.
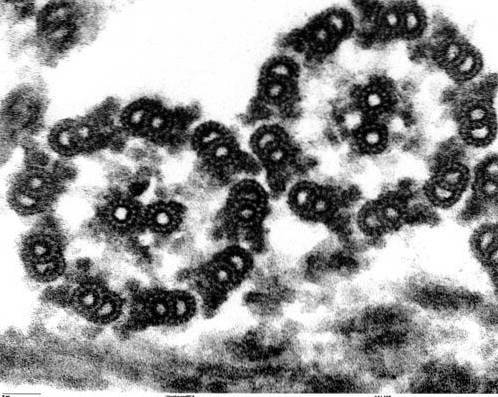
Axonemstrukturen hos cilia och flagella är mycket konservativ i alla eukaryota organismer, från mikroalger Chlamydomonas till plågan av den mänskliga spermierna.
Artikelindex
Axonemen hos den stora majoriteten av cilia och flagella har en konfiguration som kallas "9 + 2", det vill säga nio par perifera mikrotubuli som omger ett centralt par.
Mikrotubuli i varje par är olika i storlek och sammansättning, förutom det centrala paret, som presenterar båda mikrotubuli likartade. Dessa rör är stabila strukturer som kan motstå brott..
Mikrotubuli har polaritet och alla har samma arrangemang, med "+" -änden placerad mot toppen och "-" -änden basalt..
Som vi redan har påpekat är axonemets struktur av typ 9 + 2. Mikrotubuli är långa cylindriska strukturer, som består av protofilament. Protofilament består i sin tur av proteinunderenheter som kallas alfa tubulin och beta tubulin..
Varje protofilament har en alfa tubulin enhet i ena änden, medan den andra änden har en beta tubulin enhet. Slutet med beta-tubulinterminalen kallas "+" -änden, den andra änden skulle vara "-" -änden. Alla protofilament av samma mikrotubuli är orienterade med samma polaritet.
Mikrotubuli innehåller, förutom tubuliner, proteiner som kallas mikrotubuli-relaterade proteiner (MAP). Av varje par perifera mikrotubuli består den minsta (mikrotubuli A) av 13 protofilament.
Mikrotubuli B har bara 10 protofilament, men det är större än mikrotubuli A. Det centrala paret av mikrotubuli har samma storlek och var och en av dem består av 13 protofilament.
Detta centrala par mikrotubuli är inneslutet av det centrala höljet, protein i naturen, som kommer att ansluta till de perifera A-mikrotubuli med hjälp av de radiella strålarna. Å andra sidan är mikrotubuli A och B i varje par sammanfogade av ett protein som kallas nexin..
Mikrotubuli En del också ett par armar bildade av ett protein som kallas dynein. Detta protein är ansvarigt för att använda den energi som finns i ATP för att uppnå rörelse av cilia och flagella.
Externt täcks axonemet av ett ciliärt eller flagellärt membran som har samma struktur och sammansättning som plasmamembranet i cellen..
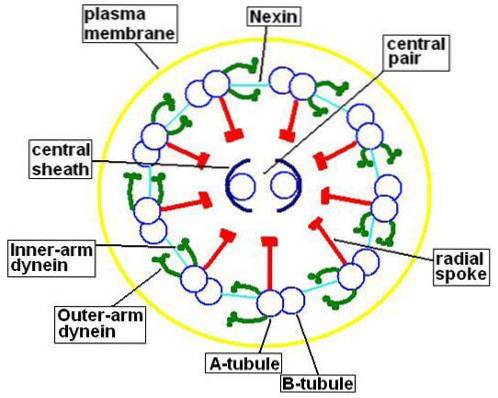
Även om axonemets "9 + 2" -komposition är mycket konserverad i de flesta eukaryota ciliated och / eller flagellerade celler, finns det några undantag från denna modell..
I spermierna hos vissa arter förloras det centrala paret av mikrotubuli, vilket resulterar i en "9 + 0" -konfiguration. Flagellrörelsen i dessa spermier tycks inte variera mycket från den som observerats i axonemer med normal konfiguration, för vilken man tror att dessa mikrotubuli inte spelar en viktig roll i rörelsen..
Detta axonemmönster har observerats i spermierna hos arter som fiskar Lycondontis och av annelider av släktet Myzostomum.
En annan konfiguration som observeras i axonemes är konfigurationen “9 + 1”. I det här fallet är en enda central mikrotubuli närvarande snarare än ett par. I sådana fall modifieras den centrala mikrotubuli omfattande och presenterar flera koncentriska väggar.
Detta axonemmönster har observerats i manliga könsceller hos vissa arter av plattmaskar. Hos dessa arter upprepas dock inte detta axonemmönster i andra flagellerade eller cilierade celler hos organismer..
Studier av flagellorörelse har visat att flagellaböjning sker utan sammandragning eller förkortning av axonemens mikrotubuli. På grund av detta har cytologen Peter Satir föreslagit en modell av flagellrörelse baserad på förskjutning av mikrotubuli..
Enligt denna modell uppnås rörelse tack vare förskjutningen av en mikrotubuli från varje par på sin partner. Detta mönster liknar glidningen av myosinkedjor på aktin under muskelsammandragning. Rörelse sker i närvaro av ATP.
Dyneinarmarna är förankrade i mikrotubuli A i varje par, med ändarna riktade mot mikrotubuli B. Vid början av rörelse vidhäftar dyneinarmarna vid bindningsstället på mikrotubuli B. Därefter sker en förändring i konfigurationen av dyneinet driver mikrorör B nedåt.
Nexin håller båda mikrotubuli nära varandra. Därefter separeras dyneinarmarna från mikrotubuli B. Det kommer sedan att återförenas med processen. Denna glidning sker växelvis mellan ena sidan av axonemet och den andra..
Denna växlande förskjutning på ena sidan av axonemet gör att cilium, eller flagellum, först böjs åt ena sidan och sedan till motsatt sida. Fördelen med Satirs flagellära rörelsemodell är att den skulle förklara appendixens rörelse oberoende av axonemkonfigurationen för axonem-mikrotubuli.
Det finns flera genetiska mutationer som kan orsaka onormal utveckling av axonemet. Dessa abnormiteter kan bland annat vara bristen på en av dyneinarmarna, antingen inre eller yttre, i de centrala mikrotubuli eller radiella strålar..
I dessa fall utvecklas ett syndrom som kallas Kartageners syndrom, där människor som lider av det är infertila eftersom spermierna inte kan röra sig.
Dessa patienter utvecklar också inälvor i en inverterad position i förhållande till den normala positionen; till exempel hjärtat på höger sida av kroppen och levern till vänster. Detta tillstånd är känt som situs inversus.
De med Kartageners syndrom är också benägna att andnings- och sinusinfektioner.
En annan sjukdom relaterad till onormal utveckling av axonemet är polycystisk njursjukdom. I detta utvecklas flera cystor i njurarna som slutligen förstör njurarna. Denna sjukdom beror på en mutation i generna som kodar för proteiner som kallas polycystiner.


Ingen har kommenterat den här artikeln än.