De centrioles De är cylindriska cellulära strukturer som består av kluster av mikrotubuli. De består av proteinet tubulin, som finns i de flesta eukaryota celler.
Ett tillhörande par centrioler, omgivna av en formlös massa av tätt material som kallas pericentriolar material (PCM), utgör en struktur som kallas centrosomen..
Centrioles funktion är att styra sammansättningen av mikrotubuli, delta i cellorganisation (position av kärnan och cellens rumsliga arrangemang), bildande och funktion av flagella och cilia (ciliogenes) och celldelning (mitos och meios).
Centrioler finns i cellulära strukturer som kallas centrosomer i djurceller och saknas i växtceller..
Defekter i strukturen eller antalet centrioler i varje cell kan ha avsevärda konsekvenser för en organisms fysiologi, vilket ger förändringar i responsen på stress under inflammation, manlig infertilitet, neurodegenerativa sjukdomar och tumörbildning, bland andra..
En centriole är en cylindrisk struktur. Ett par associerade centrioler, omgivna av en formlös massa av tätt material (kallat "pericentriolar material" eller PCM), bildar en sammansatt struktur som kallas "centrosome"..
De ansågs oviktiga förrän för några år sedan, då man drog slutsatsen att de var de viktigaste organellerna vid ledning av celldelning och duplicering (mitos) i eukaryota celler (främst hos människor och andra djur).
Den sista gemensamma förfadern för allt liv på jorden var en enda cell och den sista gemensamma förfadern för alla eukaryoter var en ciliated cell med centrioler.
Varje organism består av en grupp interagerande celler. Organismer innehåller organ, organ består av vävnader, vävnader består av celler och celler består av molekyler.
Alla celler använder samma molekylära "byggstenar", liknande metoder för att lagra, underhålla och uttrycka genetisk information och liknande processer för energimetabolism, molekylär transport, signalering, utveckling och struktur..
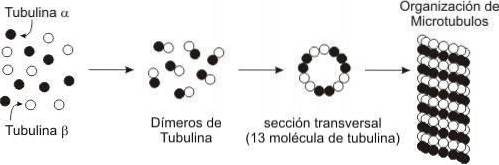
I de tidiga dagarna av elektronmikroskopi observerade cellbiologer långa tubuli i cytoplasman som de kallade mikrotubuli..
Morfologiskt liknande mikrotubuli observerades bilda fibrerna i den mitotiska spindeln, som komponenter i nervcellerna och som strukturella element i cilia och flagella..
Noggrann undersökning av de enskilda mikrotubuli indikerade att de alla bestod av 13 längsgående enheter (nu kallade protofilament) bestående av ett huvudprotein (bestående av en nära besläktad a-tubulin- och β-tubulinunderenhet) och flera proteiner associerade med mikrotubuli. (Kartor).
Förutom deras funktioner i andra celler är mikrotubuli väsentliga för neurons tillväxt, morfologi, migration och polaritet, liksom för utveckling, underhåll och överlevnad och för ett effektivt nervsystem.

Betydelsen av en känslig interaktion mellan komponenter i cytoskelettet (mikrotubuli, aktinfilament, mellanliggande filament och septiner) återspeglas i flera humana neurodegenerativa störningar relaterade till onormal mikrotubulusdynamik, inklusive Parkinsons sjukdom och Alzheimers sjukdom..
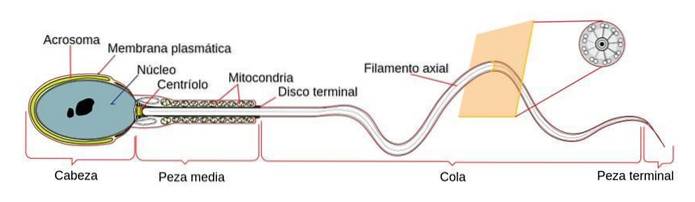
Cilia och flagella är organeller som finns på ytan av de flesta eukaryota celler. De består huvudsakligen av mikrotubuli och membran.
Spermamotilitet beror på mobila cytoskelettelement som finns i svansen, så kallade axonemer. Axonemernas struktur består av 9 grupper om 2 mikrotubuli vardera, molekylmotorer (dyneiner) och deras regulatoriska strukturer.
Centrioler spelar en central roll i ciliogenes och cellcykelprogression. Centriole mognad producerar en förändring i funktion, som leder från celldelning till ciliumbildning.
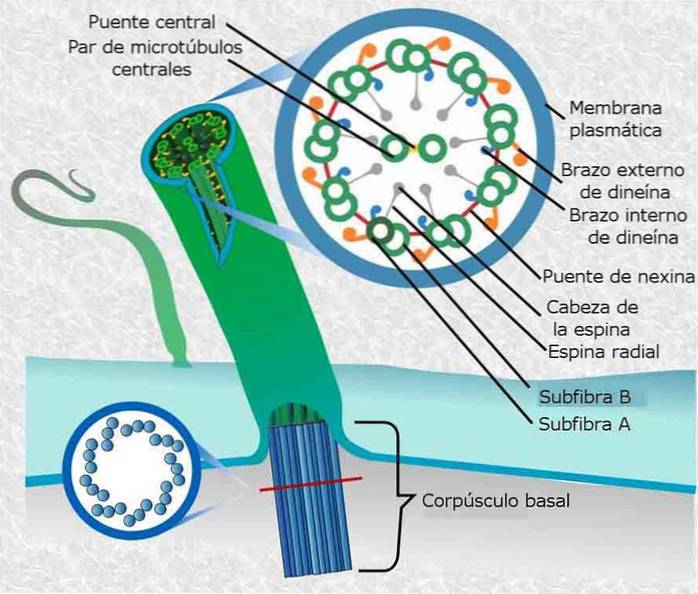
Defekter i axonemets eller cilias struktur eller funktion orsakar flera störningar hos människor som kallas ciliopatier. Dessa sjukdomar påverkar olika vävnader, inklusive ögon, njurar, hjärna, lungor och spermierörlighet (vilket ofta leder till manlig infertilitet)..
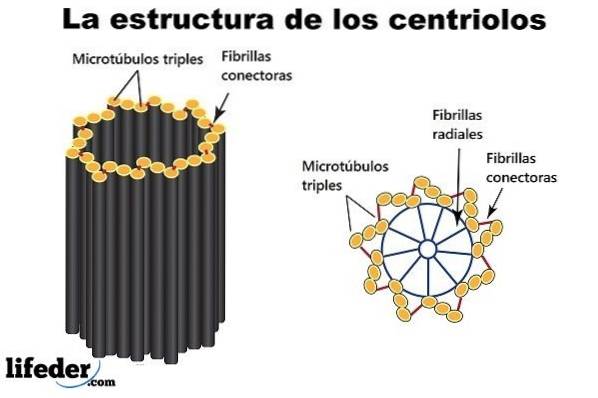
Nio tripletter av mikrotubuli anordnade runt en omkrets (bildar en kort ihålig cylinder) är ”byggstenarna” och huvudstrukturen för en centriole..
Under många år ignorerades centriolernas struktur och funktion trots att centrosomen vid 1880-talet hade visualiserats genom ljusmikroskopi..
Theodor Boveri publicerade ett banbrytande arbete 1888 och beskriver ursprunget till centrosomen från spermier efter befruktning. I sin korta kommunikation 1887 skrev Boveri att:
”Centrosomen representerar cellens dynamiska centrum; Dess uppdelning skapar centrum för de bildade dottercellerna, runt vilka alla andra cellulära komponenter är organiserade symmetriskt ... Centrosomen är cellens sanna delningsorgan, den förmedlar kärn- och celldelning ”(Scheer, 2014: 1). [Författarens översättning].
Strax efter mitten av 1900-talet, med utvecklingen av elektronmikroskopi, studerades och förklarades beteendet hos centrioler av Paul Schafer.
Tyvärr ignorerades detta arbete till stor del eftersom forskare började fokusera på resultaten från Watson och Krick på DNA..
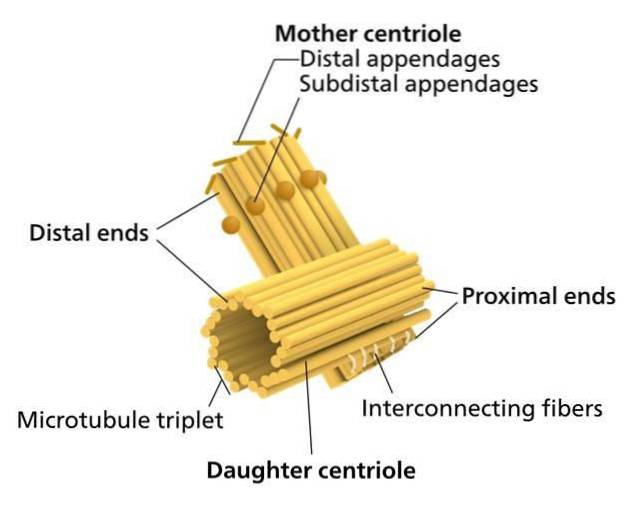
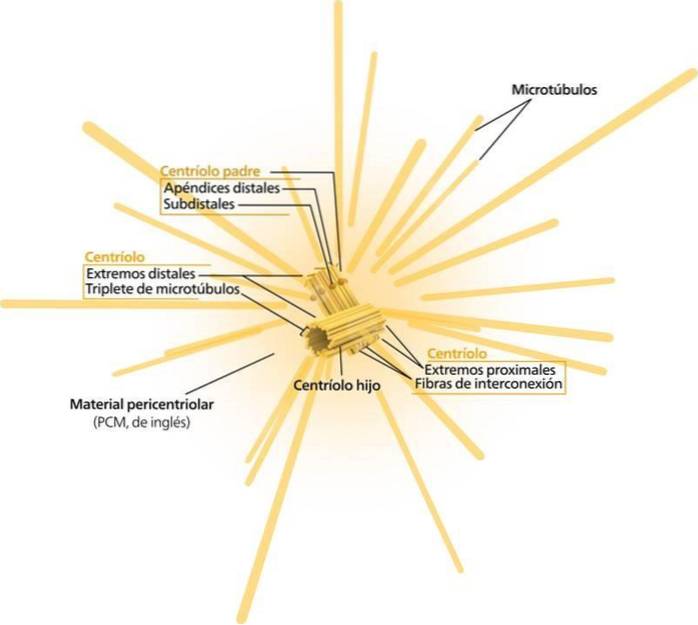
Ett par centrioler, som ligger intill kärnan och vinkelrätt mot varandra, är "en centrosom". En av centriolerna är känd som "far" (eller mor). Den andra är känd som "sonen" (eller dottern; den är något kortare och har basen fäst vid moderns bas).
De proximala ändarna (vid anslutningen av de två centriolerna) är nedsänkta i ett protein "moln" (kanske upp till 300 eller mer) känt som mikrotubuliorganiseringscentrum (MTOC), eftersom det ger det protein som är nödvändigt för konstruktion av mikrotubuli..
MTOC är också känt som "pericentriolar material" och det är negativt laddat. Omvänt är de distala ändarna (bort från anslutningen av de två centriolerna) positivt laddade.
Paret av centrioler, tillsammans med den omgivande MTOC, är kända som "centrosome".
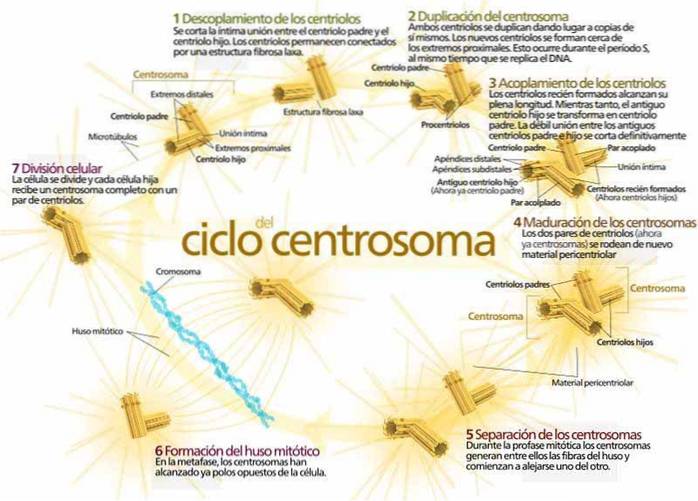
När centriolerna börjar dupliceras, separerar fadern och sonen något och sedan börjar varje centriole att bilda en ny centriole vid sin bas: fadern med en ny son och sonen med en ny son till sig (ett ”barnbarn”) ..
Medan centriolduplicering inträffar dupliceras och separerar kärn-DNA också. Det vill säga, aktuell forskning visar att centriolduplicering och DNA-separation på något sätt är kopplade..
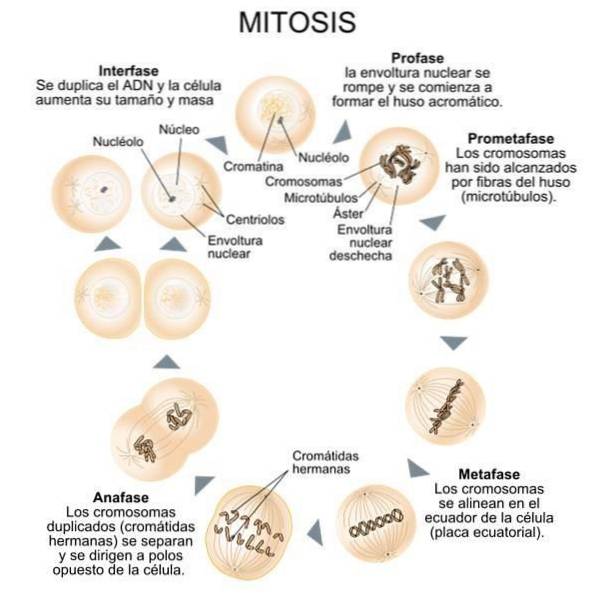
Den mitotiska processen beskrivs ofta i termer av en initiatorfas, känd som "gränssnitt", följt av fyra utvecklingsfaser..
Under interfasen dupliceras centriolerna och separeras i två par (ett av dessa par börjar röra sig mot motsatt sida av kärnan) och DNA delas..
Efter duplicering av centriolerna sträcker sig mikrotubuli i centriolerna och inriktar sig längs kärnans huvudaxel och bildar den "mitotiska spindeln".
I den första av de fyra utvecklingsfaserna (fas I eller "profas") kondenseras kromosomerna och rör sig närmare varandra och kärnmembranet börjar försvagas och lösas upp. Samtidigt bildas den mitotiska spindeln med paren av centrioler som nu är placerade i spindelns ändar..
I den andra fasen (fas II eller "metafas") inriktas kromosomkedjorna med mitotispindelns axel.
I den tredje fasen (fas III eller “anafas”) delar sig de kromosomala kedjorna och rör sig mot motsatta ändar av den nu långsträckta mitotiska spindeln..
Slutligen, i den fjärde fasen (fas IV eller “telofas”), bildas nya kärnmembran runt de separerade kromosomerna, den mitotiska spindeln faller sönder och cellseparationen börjar slutföras med hälften av cytoplasman som går med varje ny kärna..
I vardera änden av den mitotiska spindeln utövar paren av centrioler ett viktigt inflytande (tydligen relaterat till de krafter som utövas av de elektromagnetiska fälten som genereras av de negativa och positiva laddningarna i dess proximala och distala ändar) under hela celldelningsprocessen..
Exponering för stress påverkar funktion, kvalitet och livslängd för en organism. Den stress som genereras, till exempel av en infektion, kan leda till inflammation i de infekterade vävnaderna, vilket aktiverar immunsvaret i kroppen. Detta svar skyddar den drabbade organismen och eliminerar patogenen.
Många aspekter av immunsystemets funktion är välkända. De molekylära, strukturella och fysiologiska händelserna där centrosomen är involverad är dock fortfarande en gåta..
Nya studier har upptäckt oväntade dynamiska förändringar i strukturen, platsen och funktionen hos centrosomen under olika stressrelaterade förhållanden. Till exempel, efter att ha imiterat villkoren för en infektion, har en ökning av PCM- och mikrotubulusproduktion påträffats i interfasceller..
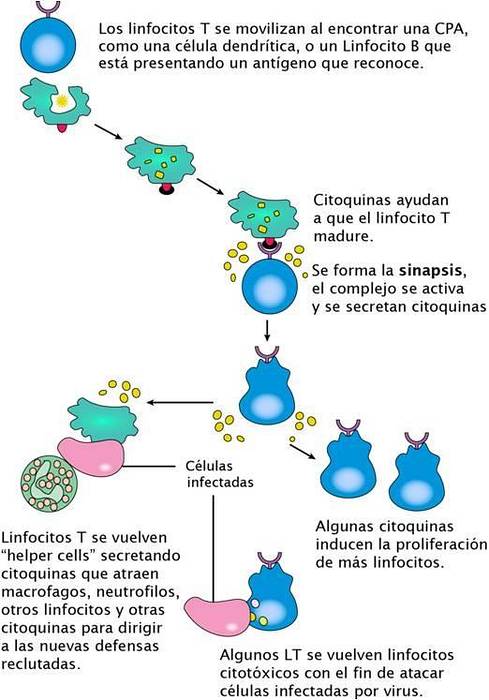
Centrosomen spelar en mycket viktig roll i strukturen och funktionen hos den immunologiska synapsen (SI). Denna struktur bildas av specialiserade interaktioner mellan en T-cell och en antigenpresenterande cell (APC). Denna cell-cell-interaktion initierar centrosom-migrering mot SI och dess efterföljande koppling till plasmamembranet..
Centrosomkopplingen i SI liknar den som observerades under ciliogenes. I det här fallet initierar det emellertid inte sammansättningen av cilia utan deltar snarare i organisationen av SI och utsöndringen av cytotoxiska vesiklar för att lysera målcellerna och blir ett nyckelorgan vid aktivering av T-celler..
Centrosomen är målet för ”molekylära chaperoner” (uppsättning proteiner vars funktion är att hjälpa vikning, montering och cellulär transport av andra proteiner) som ger skydd mot exponering för värmechock och stress..
Stressfaktorer som påverkar centrosomen inkluderar DNA-skador och värme (som det som upplevs av celler från feberpatienter). DNA-skada initierar DNA-reparationsvägar, vilket kan påverka centrosomfunktionen och proteinsammansättningen.
Spänningen som genereras av värme orsakar modifiering av centriole-strukturen, störningen av centrosomen och fullständig inaktivering av dess förmåga att bilda mikrotubuli, vilket förändrar bildningen av den mitotiska spindeln och förhindrar mitos.
Störning av centrosomefunktionen under feber kan vara en adaptiv reaktion för att inaktivera spindelstolparna och förhindra onormal DNA-uppdelning under mitos, särskilt med tanke på den potentiella dysfunktionen hos flera proteiner efter värmeinducerad denaturering..
Det kan också ge cellen extra tid att återvinna sin pool av funktionella proteiner innan celldelningen startas om..
En annan konsekvens av inaktiveringen av centrosomen under feber är dess oförmåga att överföra till SI för att organisera den och delta i utsöndringen av cytotoxiska vesiklar..
Utvecklingen av centriolen är en ganska komplex process och även om en serie reglerande proteiner deltar i den kan olika typer av fel uppstå.
Om det finns en obalans i förhållandet mellan proteiner kan dottercentriolen vara defekt, dess geometri kan förvrängas, axlarna i ett par kan avvika från vinkelrätt, flera dottercentrioler kan utvecklas, dottercentriolen kan nå full längd före tiden, eller frikopplingen av paren kan försenas.
När det finns fel eller fel duplicering av centrioler (med geometriska defekter och / eller multipel duplicering) förändras DNA-replikering, kromosomal instabilitet (CIN) inträffar.
På samma sätt leder centrosomfel (till exempel en förstorad eller förstorad centrosom) till CIN och främjar utvecklingen av flera dottercentrioler..
Dessa utvecklingsfel genererar skador på celler som till och med kan leda till maligna celler..
Tack vare ingripandet av reglerande proteiner, när abnormiteter upptäcks i utvecklingen av centriolerna och / eller centrosomen, kan cellerna genomföra självkorrigering av abnormiteterna.
Om självkorrigering av anomali inte uppnås kan dock onormala centrioler eller med flera barn ("supernumerära centrioler") leda till generering av tumörer ("tumörgenes") eller celldöd..
Supernumerära centrioler tenderar att sammanfalla, vilket leder till gruppering av centrosomen ("centrosomförstärkning", karakteristisk för cancerceller), vilket förändrar cellpolariteten och den normala utvecklingen av mitos, vilket resulterar i uppkomsten av tumörer..
Celler med supernumerära centrioler kännetecknas av ett överskott av pericentriolar material, avbrott i den cylindriska strukturen eller överdriven längd av centriolerna och centriolerna som inte är vinkelräta eller dåligt placerade..
Det har föreslagits att kluster av centrioler eller centrosomer i cancerceller skulle kunna tjäna som en "biomarkör" vid användning av terapeutiska och bildande medel, såsom superparamagnetiska nanopartiklar..



Ingen har kommenterat den här artikeln än.