De oxidativ fosforylering är en process där ATP-molekyler syntetiseras från ADP och Pi (oorganiskt fosfat). Denna mekanism utförs av bakterier och eukaryota celler. I eukaryota celler sker fosforylering i den mitokondriella matrisen för icke-fotosyntetiska celler.
ATP-produktion drivs av överföring av elektroner från koenzymerna NADH eller FADHtvå till Otvå. Denna process representerar den högsta energiproduktionen i cellen och härrör från nedbrytningen av kolhydrater och fetter..
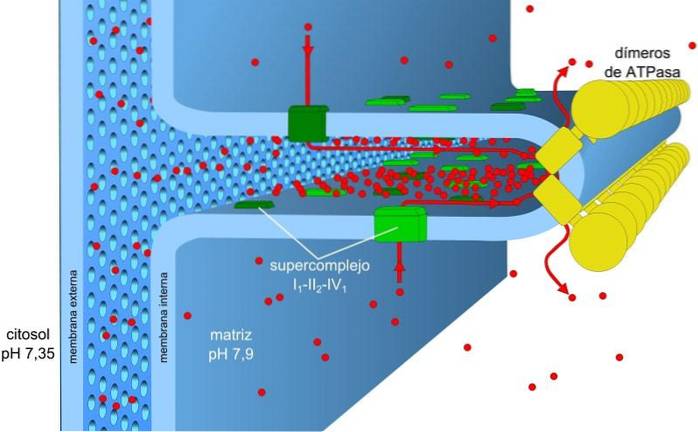
Den energi som lagras i laddnings- och pH-gradienterna, även känd som protonmotivkraften, gör att denna process kan äga rum. Protongradienten som genereras får den yttre delen av membranet att ha en positiv laddning på grund av koncentrationen av protoner (H+) och den mitokondriella matrisen är negativ.
Artikelindex
Processerna för elektrontransport och oxidativ fosforylering är associerade med ett membran. I prokaryoter utförs dessa mekanismer genom plasmamembranet. I eukaryota celler associeras de med mitokondriellt membran.
Antalet mitokondrier som finns i celler varierar beroende på celltyp. Till exempel, hos däggdjur saknar erytrocyter dessa organeller, medan andra celltyper, såsom muskelceller, kan ha upp till miljoner av dem..
Det mitokondriella membranet består av ett enkelt yttre membran, ett något mer komplext inre membran, och däremellan intermembranutrymmet, där många ATP-beroende enzymer finns.
Det yttre membranet innehåller ett protein som kallas porin som bildar kanalerna för enkel diffusion av små molekyler. Detta membran är ansvarigt för att upprätthålla mitokondriernas struktur och form.
Det inre membranet har högre densitet och är rikt på proteiner. Det är också ogenomträngligt för molekyler och joner, så för att korsa det behöver de intermembranproteiner för att transportera dem..
Inuti matrisen sträcker sig vikningarna på det inre membranet och bildar åsar som gör det möjligt att ha ett stort område i en liten volym.
Mitokondrion anses vara den centrala producenten av cellulär energi. Den innehåller de enzymer som är involverade i processerna för citronsyracykeln, fettsyraoxidation och redoxenzymer och proteiner för elektrontransport och ADP-fosforylering.
Protonkoncentrationsgradienten (pH-gradienten) och laddningsgradienten eller den elektriska potentialen i mitokondriernas inre membran är ansvariga för protonens drivkraft. Den inre membranets låga permeabilitet för joner (andra än H+) tillåter mitokondrier att ha en stabil spänningsgradient.
Elektronisk transport, protonpumpning och ATP-erhållning sker samtidigt i mitokondrierna, tack vare protonmotivkraften. PH-gradienten upprätthåller sura förhållanden i intermembranet och den mitokondriella matrisen med alkaliska förhållanden.
För varannan elektron som överförs till Otvå cirka 10 protoner pumpas genom membranet, vilket skapar en elektrokemisk gradient. Den energi som frigörs i denna process produceras gradvis genom att elektroner passerar genom transportkedjan.
Energin som frigörs under oxidationsreduktionsreaktionerna av NADH och FADHtvå är avsevärt högt (cirka 53 kcal / mol för varje elektronpar), så för att kunna användas vid tillverkning av ATP-molekyler, måste det produceras gradvis med passage av elektroner genom transportörer.
Dessa är organiserade i fyra komplex belägna på det inre mitokondriella membranet. Kopplingen av dessa reaktioner till syntesen av ATP utförs i ett femte komplex.
NADH överför ett par elektroner som går in i komplex I i elektrontransportkedjan. Elektroner överförs till flavinmononukleotiden och sedan till ubiquinon (koenzym Q) genom en järn-svaveltransportör. Denna process frigör en stor mängd energi (16,6 kcal / mol).
Ubiquinone transporterar elektroner över membranet till komplex III. I detta komplex passerar elektronerna genom cytokromer b och c1 tack vare en järn-svaveltransportör.
Elektroner passerar från komplex III till komplex IV (cytokrom c oxidas), överförs en efter en i cytokrom c (perifert membranprotein). I komplex IV passerar elektronerna genom ett par kopparjoner (Cutilltvå+följt av cytokrom ctill, sedan till ett annat par kopparjoner (Cubtvå+) och från detta till cytokrom a3.
Slutligen överförs elektronerna till Otvå som är den sista acceptorn och bildar en vattenmolekyl (HtvåO) för varje mottagen elektron. Elektronens passage från komplex IV till Otvå Det genererar också en stor mängd fri energi (25,8 kcal / mol).
Komplex II (succinat CoQ-reduktas) tar emot ett par elektroner från citronsyracykeln genom oxidation av en succinatmolekyl för att fumera. Dessa elektroner överförs till FAD och passerar sedan genom en järn-svavelgrupp till ubiquinon. Från detta koenzym passerar de till komplex III och följer den tidigare beskrivna vägen.
Energin som frigörs vid elektronöverföringsreaktionen till FAD räcker inte för att driva protonerna genom membranet, så ingen protonkraft genereras i detta steg i kedjan, och följaktligen ger FADH mindre H+ att NADH.
Den energi som genereras i den elektrontransportprocess som beskrivits tidigare måste kunna användas för produktion av ATP, en reaktion katalyserad av enzymet ATP-syntas eller komplex V. Bevarandet av denna energi är känd som energikoppling och mekanismen har svårt att karakterisera.
Flera hypoteser har beskrivits för att beskriva denna energitransduktion. Det bäst accepterade är den kemiosmotiska kopplingshypotesen, som beskrivs nedan..
Denna mekanism föreslår att energin som används för ATP-syntes kommer från en protongradient i cellmembran. Denna process ingriper i mitokondrier, kloroplaster och bakterier och är kopplad till transport av elektroner.
Komplexen I och IV i elektrontransport fungerar som protonpumpar. Dessa genomgår konforma förändringar som gör det möjligt för dem att pumpa protoner i intermembranutrymmet. I komplex IV pumpas två protoner ut ur membranet för varje elektronpar, och ytterligare två finns kvar i matrisen och bildar HtvåELLER.
Ubiquinone i komplex III accepterar protoner från komplexen I och II och släpper ut dem på utsidan av membranet. Komplexen I och III medger vardera passage av fyra protoner för varje par transporterade elektroner.
Den mitokondriella matrisen har en låg koncentration av protoner och en negativ elektrisk potential, medan det intermembrana utrymmet presenterar de inversa förhållandena. Flödet av protoner genom detta membran representerar den elektrokemiska gradienten som lagrar den nödvändiga energin (± 5 kcal / mol per proton) för syntesen av ATP..
Enzymet ATP-syntetas är det femte komplexet som är involverat i oxidativ fosforylering. Det är ansvarigt för att utnyttja den elektrokemiska gradientens energi för att bilda ATP.
Detta transmembranprotein består av två komponenter: F0 och F1. Komponent F0 tillåter återgång av protoner till den mitokondriella matrisen som fungerar som en kanal och F1 katalyserar syntesen av ATP med ADP och P.i, använda energin i nämnda retur.
ATP-syntesprocessen kräver en strukturell förändring av F1 och montering av komponenter F0 och F1. Protontranslokering genom F0 orsakar konformationsförändringar i tre underenheter av F1, låter den fungera som en rotationsmotor som styr bildandet av ATP.
Den underenhet som ansvarar för bindningen av ADP med Pi den går från ett svagt tillstånd (L) till ett aktivt (T). När ATP bildas går en andra subenhet i ett öppet tillstånd (O) som möjliggör frisättning av denna molekyl. Efter att ATP släppts går denna underenhet från öppet tillstånd till inaktivt tillstånd (L).
ADP- och P-molekyleri binda till en underenhet som har gått från ett O-tillstånd till ett L-tillstånd.
Elektrontransportkedjan och fosforylering producerar ATP-molekyler. Oxidationen av NADH producerar cirka 52,12 kcal / mol (218 kJ / mol) fri energi.
Den totala reaktionen för oxidation av NADH är:
NADH + 1⁄2 Otvå +H+ ↔ HtvåO + NAD+
Överföring av elektroner från NADH och FADHtvå inträffar genom olika komplex, så att den fria energiförändringen ΔG ° kan delas upp i mindre energi "paket", som är kopplade till ATP-syntes.
Oxidationen av en NADH-molekyl genererar syntesen av tre ATP-molekyler. Medan oxidationen av en FADH-molekyltvå par till syntesen av två ATP: er.
Dessa koenzymer kommer från glykolys- och citronsyracykelprocesserna. För varje glukosmolekyl som bryts ned producerar de 36 eller 38 molekyler ATP, beroende på var cellerna befinner sig. 36 ATP produceras i hjärnan och skelettmuskeln, medan 38 ATP produceras i muskelvävnad..
Alla organismer, encelliga och flercelliga, behöver minimalt med energi i sina celler för att utföra processerna inom dem och i sin tur upprätthålla vitala funktioner i hela organismen..
Metaboliska processer kräver energi att äga rum. Det mesta av den användbara energin erhålls genom nedbrytning av kolhydrater och fetter. Denna energi härrör från oxidativ fosforyleringsprocess.
ATP-användningshastigheten i celler styr dess syntes, och i sin tur, på grund av kopplingen av oxidativ fosforylering med elektrontransportkedjan, reglerar den i allmänhet också hastigheten för elektrontransport..
Oxidativ fosforylering har en strikt kontroll som säkerställer att ATP inte genereras snabbare än det konsumeras. Det finns vissa steg i elektrontransport och kopplad fosforylering som reglerar energiproduktionens hastighet..
De viktigaste vägarna för energiproduktion (cellulär ATP) är glykolys, citronsyracykeln och oxidativ fosforylering. Samordnad kontroll av dessa tre processer reglerar ATP-syntes.
Kontrollen av fosforylering av ATP: s massverkningsförhållande beror på elektronernas exakta bidrag i transportkedjan. Detta beror i sin tur på [NADH] / [NAD+] som hålls förhöjd av glykolysverkan och citronsyracykeln.
Denna samordnade kontroll utförs genom reglering av glykolyskontrollpunkterna (PFK inhiberad av citrat) och citronsyracykeln (Pyruvatdehydrogenas, citrat tapeas, isocitratdehydrogenas och a-ketoglutarat dehydrogenas)..
Komplex IV (cytokrom c oxidas) är ett enzym som regleras av ett av dess substrat, det vill säga dess aktivitet styrs av reducerad cytokrom c (ctvå+), vilket i sin tur är i jämvikt med koncentrationsförhållandet mellan [NADH] / [NAD+] och massverkningsförhållandet mellan [ATP] / [ADP] + [Pi].
Ju högre förhållande [NADH] / [NAD+] och sänk [ATP] / [ADP] + [Pi], ju högre koncentration av cytokrom [ctvå+] och ju större aktivitet av komplex IV. Detta tolkas till exempel om vi jämför organismer med olika aktiviteter för vila och hög aktivitet.
Hos en person med hög fysisk aktivitet, konsumtionen av ATP och därmed dess hydrolys till ADP + Pi kommer att vara mycket hög, vilket genererar en skillnad i massverkningsförhållandet som orsakar en ökning av [ctvå+] och därför en ökning av ATP-syntes. Hos en individ i vila inträffar den omvända situationen.
I slutändan ökar hastigheten för oxidativ fosforylering med koncentrationen av ADP i mitokondrierna. Denna koncentration beror på ADP-ATP-translokatorerna som ansvarar för transporten av adeninnukleotider och Pi från cytosolen till den mitokondriella matrisen.
Oxidativ fosforylering påverkas av vissa kemiska agenser som gör att elektrontransport fortsätter utan ADP-fosforylering, vilket kopplar bort energiproduktion och konservering.
Dessa medel stimulerar mitokondriernas syreförbrukningshastighet i frånvaro av ADP, vilket också orsakar en ökning av ATP-hydrolys. De agerar genom att eliminera en mellanhand eller bryta ett energitillstånd i elektrontransportkedjan.
2,4-dinitrofenol, en svag syra som passerar genom mitokondriella membran, är ansvarig för att skingra protongradienten, eftersom de binder till dem på den sura sidan och släpper ut dem på basen.
Denna förening användes som ett "bantningspiller" eftersom det visade sig producera en ökning av andningen, därför en ökning av ämnesomsättningshastigheten och tillhörande viktminskning. Det visades dock att dess negativa effekt till och med kan orsaka dödsfall.
Förlusten av protongradienten producerar värme. Celler i brun fettvävnad använder hormonstyrd frikoppling för att producera värme. Vilande däggdjur och nyfödda som saknar hår består av denna vävnad som fungerar som en slags termisk filt..
Föreningar eller hämmande medel förhindrar båda konsumtionen av Otvå (elektrontransport) såsom tillhörande oxidativ fosforylering. Dessa medel förhindrar bildandet av ATP genom att använda den energi som produceras vid elektronisk transport. Därför stannar transportkedjan eftersom nämnda energiförbrukning inte är tillgänglig..
Det antibiotiska oligomycinet fungerar som en fosforyleringshämmare i många bakterier, vilket förhindrar stimulering av ADP till ATP-syntes.
Det finns också jonoformedel som bildar fettlösliga komplex med katjoner såsom K+ och Na+, och de passerar genom mitokondriellt membran med dessa katjoner. Mitokondrierna använder sedan den energi som produceras vid elektrontransport för att pumpa katjoner istället för att syntetisera ATP..



Ingen har kommenterat den här artikeln än.