De ontogeni det är den process genom vilken en individs utveckling sker. Fenomenet börjar med befruktning och sträcker sig till åldrande av organiska varelser. Fältet biologi som ansvarar för att studera ontogeni är utvecklingsbiologi.
I denna process inträffar ”översättningen” av genotypen - all genetisk information från en biologisk enhet - till den fenotyp som vi kan observera. Den mest dramatiska omvandlingen sker i de tidiga utvecklingsstadierna, med omvandlingen av en cell till en fullständig individ.
Idag är sammanslagningen av utvecklingsbiologi och evolutionsteori, känd som evo-devo, en mycket populär kunskap som växer med stormsteg. Detta nya fält syftar till att förklara utvecklingen av den enorma mångfalden av morfologier som levande organismer uppvisar..
Artikelindex
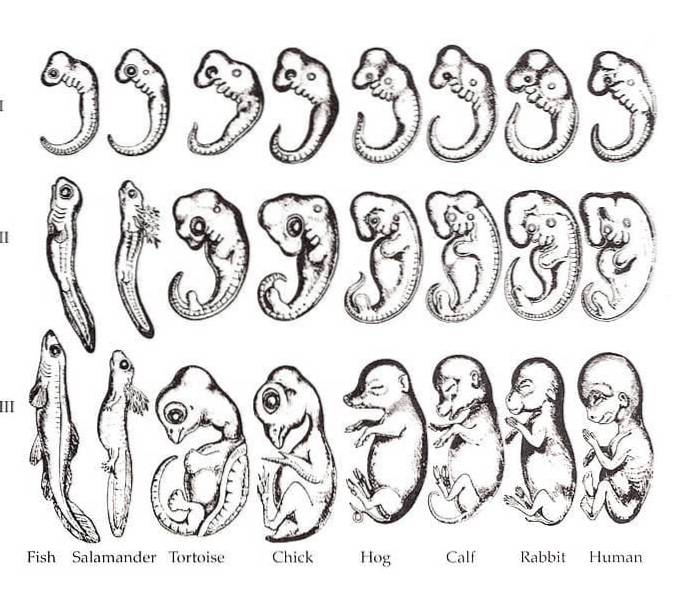
Förhållandet mellan ontogeni och fylogeni var en dominerande uppfattning under hela 2000-talet. Det är allmänt känt att olika arter av organismer är mycket mer lika varandra i sina embryoniska stadier än de är i vuxna former. År 1828 märkte Karl Ernst von Baer detta mönster i sybphylum Vertebrata.
Baer noterade att det i olika arter av tetrapoder finns vissa likheter i embryot, såsom gälar, notokord, segmentering och fenformade lemmar..
Dessa bildas före de typiska egenskaperna som gör att gruppen i fråga kan diagnostiseras i en mer specifik hierarkisk klassificeringsordning.
Denna idé omformulerades av den berömda - och en av Charles Darwins mest passionerade anhängare - den tyskfödda biologen Ernst Haeckel..
Haeckel krediteras med den berömda frasen "ontogeny recapitulates fylogeny." Med andra ord föreslår rekapitulationen att utvecklingen av en organism upprepar dess evolutionära historia från vuxna former av dess förfäder..
Även om frasen är välkänd idag, var det vid mitten av 2000-talet klart att Haeckels förslag sällan uppfylls..
S. J. Gould, den berömda paleontologen och evolutionärbiologen, lade fram sina idéer om rekapitulering i det han kallade "terminaladditionsprincipen." För Gould kan rekapitulering inträffa så länge som evolutionär förändring inträffar genom successiva tillägg av stadier i slutet av en förfäders ontogeni..
På samma sätt måste det också uppfyllas att den tidliga varaktigheten för förfäders ontogeni måste förkortas när släktlinjen utvecklades.
Idag har moderna metoder lyckats motbevisa begreppet addition som föreslås av den biogenetiska lagen.
För Haeckel inträffade detta tillägg på grund av den kontinuerliga användningen av organen. Emellertid har de evolutionära konsekvenserna av användning och användning av organ kasserats.
Det är för närvarande känt att gälvbågarna i de embryonala stadierna hos däggdjur och reptiler aldrig har den form som motsvarar vuxen fisk.
Dessutom finns det variationer i tidpunkten eller tidpunkten för vissa utvecklingsstadier. I evolutionsbiologin kallas denna förändring heterokroni..
Ontogeny omfattar alla processer för utveckling av organiska varelser, som börjar med befruktning och slutar med åldrande..
Logiskt sett sker de mest dramatiska omvandlingarna i de första stegen, där en enda cell kan bilda en hel individ. Därefter kommer vi att beskriva ontogeniprocessen och betona embryonstegen.
Under processen med oogenes förbereder ett ägg (den kvinnliga könscellen, även kallad ett ägg) för befruktning och de tidiga utvecklingsstadierna. Detta sker genom att ackumulera reservmaterial för framtiden..
Äggets cytoplasma är en miljö rik på olika biomolekyler, huvudsakligen budbärar-RNA, ribosomer, överförings-RNA och andra maskiner som är nödvändiga för syntes av proteiner. Cellkärnan genomgår också betydande tillväxt.
Sperma behöver inte denna process, deras strategi är att eliminera så mycket cytoplasma som möjligt och kondensera kärnan för att bevara små dimensioner..
Händelsen som markerar början på ontogeni är befruktning, som involverar en manlig och kvinnlig könsförening, vanligtvis under sexuell reproduktion..
I fallet med extern befruktning, som förekommer i många marina organismer, utvisas båda könscellerna i vattnet och hittas slumpmässigt.
Vid befruktning återintegreras individets diploida antal och möjliggör kombinationsprocesserna mellan faderns och moderns gener.
I vissa fall är spermier inte nödvändiga för att aktivera utveckling. Men hos de flesta individer utvecklas inte embryot på rätt sätt. På samma sätt kan vissa arter reproducera genom parthenogenes, där normal embryoutveckling sker utan behov av spermier..
Däremot kräver vissa ägg aktivering av spermierna, men införlivar inte det genetiska materialet i denna manliga könsceller i embryot..
Sperma och ägg måste kännas igen korrekt så att alla händelser efter befruktning kan äga rum. Detta igenkännande förmedlas av en serie av artsspecifika proteiner. Det finns också hinder som förhindrar att ett ägg, när det befruktats, nås av en andra sperma.
Efter befruktning och aktivering av ägget inträffar de första utvecklingsstadierna. I segmentering delar sig embryot upprepade gånger för att bli en grupp celler som kallas blastomerer..
Under denna sista period inträffar ingen celltillväxt, utan endast indelningen av massan sker. I slutändan finns det hundratals eller tusentals celler som viker för blastula-tillståndet.
När embryot utvecklas får det en polaritet. Av denna anledning är det möjligt att skilja mellan växtpolen, som ligger i ena änden, och djurpolen, rik på cytoplasma. Denna axel utgör en referenspunkt för utveckling.
Beroende på mängden äggula som ägget har och fördelningen av nämnda substans kan ägget klassificeras som oligolecyter, heterolecyter, telolecyter och centrolecyter..
De förstnämnda har, som namnet antyder, en liten mängd äggula och den är mer eller mindre jämnt fördelad över ägget. Generellt är dess storlek liten. Heterolecyter har mer äggula än oligolecyter och äggula koncentreras i den vegetativa polen.
Telolecitos har en riklig mängd äggula och upptar nästan hela ägget. Slutligen har centrolecitos all äggula koncentrerad i äggets centrala region..
Blastula är en massa celler. I däggdjur kallas denna cellgrupp för en blastocyst, medan cellerna i de flesta djur är ordnade runt en central vätskekavitet, kallad blastocele..
I blastula-tillståndet har det varit möjligt att visa en stor ökning när det gäller mängden DNA. Emellertid är storleken på hela embryot inte mycket större än den ursprungliga zygoten..
Gastrulering omvandlar den sfäriska och enkla blastula till en mycket mer komplex struktur med två groddskikt. Denna process är heterogen om vi jämför djurens olika släkter. I vissa fall bildas ett andra lager utan att göra ett inre hålrum.
Öppningen till tarmen kallas blastopore. Blastopores öde är en mycket viktig egenskap för uppdelningen av två stora släkter: protostomaterna och deuterostomerna. I den första gruppen ger blastopore upphov till munnen, medan i den andra ger blastopore anus.
Således har gastrula två lager: ett yttre lager som omger blastocele, kallat ektoderm, och ett inre lager som kallas endoderm..
De flesta djur har ett tredje groddskikt, mesoderm, som ligger mellan de två skikten som nämns ovan. Mesoderm kan bildas på två sätt: cellerna uppstår från en ventral region av blastopores läpp och därifrån sprider de sig eller uppstår från den centrala regionen av archnteronväggarna..
I slutet av gastrulering täcker ektoderm embryot och mesoderm och endoderm ligger i den inre delen. Med andra ord har cellerna en annan slutposition än den där de startade..
Coelom är ett kroppshålrum som omges av mesoderm. Detta inträffar eftersom blastocele nästan fylls med mesoderm under gastrulationsprocessen..
Detta coelomatiska hålrum kan förekomma på två sätt: schizocelic eller enterocelic. Båda coeloms är dock funktionellt ekvivalenta.
Organogenes innefattar en serie processer där vart och ett av organen bildas.
De mest relevanta händelserna inkluderar migrering av vissa celler till den plats där de är nödvändiga för att bilda nämnda organ..
Under utveckling har det fastställts att epigenes fortskrider i tre steg: mönsterbildning, bestämning av kroppsposition och induktion av rätt position för extremiteterna och olika organ.
För att generera ett svar finns det vissa genprodukter, kallade morfogener (definitionen av dessa enheter är teoretisk, inte kemisk). Dessa fungerar tack vare bildandet av en differentiell gradient som ger rumslig information.
Beträffande de inblandade generna spelar homeotiska gener en grundläggande roll i utvecklingen av individer eftersom de definierar segmentens identitet.



Ingen har kommenterat den här artikeln än.