
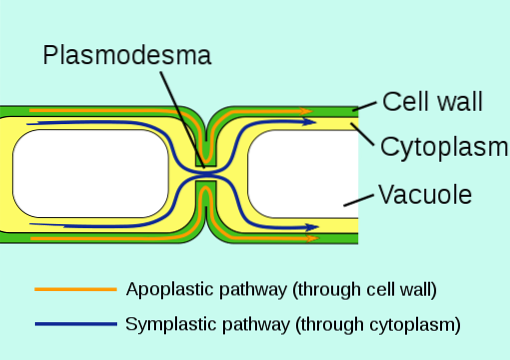
De plasmodesmer är cytosoliska anslutningar som inträffar mellan intilliggande växtceller, det vill säga de kommunicerar protoplasterna (plasmamembran och cytosol) genom cellväggen och bildar ett förenklat kontinuum.
Dessa strukturer är funktionellt analoga eller ekvivalenta med klyvkorsningar (gap korsningar) som observeras mellan cellerna i en djurvävnad och deras huvudsakliga funktion är att kommunicera cellerna med varandra och fungera som en kanal för transport av olika typer av joner och molekyler.
Plasmodesmata beskrevs för mer än 100 år sedan av Tangl och sedan dess har hundratals studier publicerats där deras funktionsmekanism, deras struktur och andra relaterade aspekter har detaljerats i detalj..
För närvarande är det känt att dessa cytosoliska "kanaler" eller "förbindelser" mellan celler är strukturer under strikta kontrollmekanismer och det har också bestämts att de huvudsakligen består av integrerade membranproteiner, chaperonproteiner och andra proteiner specialiserade på transport av ämnen..
Artikelindex
Plasmodesmata ansluter celler som tillhör samma "förenklade domän" i en växtvävnad, vilket innebär att inte alla celler i en växt är kopplade till varandra, men det finns olika specifika "regioner" i en vävnad där cellerna som finns där utbyter information permanent.
Dessa är mycket dynamiska strukturer; deras antal, struktur och funktion kan modifieras som svar på ett specifikt funktionellt krav på en vävnad.
Dessutom kan dessa kanaler brytas ned eller "förseglas" i vissa cellulära gränssnitt (utrymmet mellan två celler), vilket innebär bildandet av en förenklad "barriär" mellan cellerna i vissa växtvävnader och gynnar isoleringen av definierade regioner i en vävnad.
Vissa bibliografiska citat tyder på att plasmodesmata är strukturer lika komplexa som de så kallade kärnporekomplexen, som utför liknande funktioner men vid translokering av molekylär information från den cytosoliska miljön till det inre av kärnan..
En snabb blick på en vävnadsvävnad räcker för att verifiera att det finns flera typer av plasmodesmata..
Enligt vissa författare kan dessa klassificeras som primära och sekundära enligt det ögonblick då de bildas under en cells liv; eller så enkelt och grenat, beroende på morfologin hos kanalerna som bildas mellan cell och cell.
Oavsett vilken typ av plasmodesmus det är fråga om, är dess "strukturella arkitektur" mer eller mindre ekvivalent, eftersom det nästan alltid handlar om kanaler med en diameter som varierar mellan 20 och 50 nm, vars ingångar eller öppningar är lite smalare, utgör det som kallas en "flaskhalssträngning".
Vissa forskare har föreslagit att sådan förträngning i plasmodesmatas öppningar deltar i regleringen av ämnens flöde genom dem, det vill säga att deras utvidgning (expansion) eller förträngning (minskning i diameter) bestämmer mängden och hastigheten på flödet..
Dessa "flaskhalsar" är sammansatta av ett ämne som kallas kallos (β-1,3-glukan), och som man kan sluta sig till finns de i de områden som ligger närmast väggen till växtceller som är anslutna genom dessa kanaler..
Primär plasmodesmata bildas i "cellplattan" under cytokinese, vilket är tiden för mitos där de två dottercellerna separerar. Dessa kan dock genomgå strukturella modifieringar och ändra distribution och drift under utvecklingen av anläggningen som de tillhör..
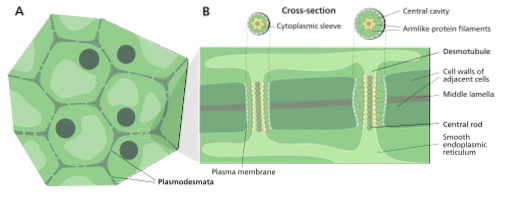
Dessa plasmodesmata är faktiskt membranmiljöer som består av porer i plasmamembranet som bildar en slags bro mellan cellväggen och ett axiellt element i det "fångade" endoplasmiska nätverket kallat desmotubuli..
En demotubule är en cylindrisk struktur med mer eller mindre 15 nm i diameter, bestående av det endoplasmiska retikulumet i en cell som är kontinuerlig med cisternerna i det endoplasmiska retikulumet i den angränsande cellen som är ansluten genom plasmodesmen..
Mellan "strängen" representerad av demotubulen och plasmamembranet som utgör det cylindriska hålrummet som är plasmodesmus finns det ett utrymme som kallas "cytoplasmatisk hylsa" (från engelska Cytoplasmatisk hylsa), vilket är genom vilket flödet av ämnen från en cell till en annan antas ske.
Dessa är de som kan bildas de novo mellan två cellväggar oberoende av cytokinese, det vill säga utan behov av en celldelningshändelse. Sekundära plasmodesmata anses ha speciella funktionella och strukturella egenskaper.
Sekundära plasmodesmata bildas genom fusion av motsatta ändar av redan existerande "halvor" av plasmodesmata, vilka vanligtvis är etablerade i områden av cellväggen som har tunnats ut. Varje fusionshalva skapar de centrala håligheterna i en plasmodesmus.
De centrala strängarna i denna typ av plasmodesm läggs senare till genom passiv "inneslutning" av endoplasmatiska retikulumrör och den resulterande morfologin liknar den primära plasmodesmata..
Experter inom området föreslår att sekundära plasmodesmata bildas i celler som genomgår omfattande tillväxtprocesser (töjning), det vill säga mellan längsgående cellväggar, för att kompensera för den progressiva "utspädningen" av antalet plasmodesmata som kan uppstå tack vare tillväxt.
Plasmodesmata representerar en av de viktigaste cellcellskommunikationsvägarna i växtvävnad. Dessa strukturer erbjuder också en kanal för elektrisk signalering, för diffusion av lipider och små lösliga molekyler och till och med för utbyte av transkriptionsfaktorer och makromolekyler såsom proteiner och nukleinsyror..
Dessa kommunikationsvägar som tillhandahålls av plasmodesmata verkar ha en väsentlig funktion i programmeringen av växtutveckling och även i samordningen av en mogen växts fysiologiska funktion..
De deltar i regleringen av frisättningen av viktiga molekyler från den fysiologiska och utvecklingsmässiga synvinkeln mot flödet (som bär saften); de ingriper i den fysiska isoleringen av vissa celler och vävnader under utveckling, varför de sägs samordna tillväxt, utveckling och försvar mot patogener.
Efter invasion av en patogen svamp är plasmodesmata också involverade, eftersom de motsvarar den huvudsakliga intracellulära eller förenklade invasionen i växtvävnader..


Ingen har kommenterat den här artikeln än.