De DNA-förpackning är en term som definierar den kontrollerade komprimeringen av DNA i cellen. I ingen cell (och faktiskt inte ens i virus) är DNA-fri, lös och i riktig lösning.
DNA är en extremt lång molekyl som också alltid interagerar med ett stort antal olika proteiner. För bearbetning, arv och kontroll av uttrycket av generna som det bär, antar DNA en viss rumslig organisation. Detta uppnås av cellen genom att strikt kontrollera varje steg av DNA-förpackning vid olika komprimeringsnivåer..
Virus har olika förpackningsstrategier för sina nukleinsyror. En av favoriterna är den som bildar kompakta spiraler. Man kan säga att virus är nukleinsyror förpackade i sina egna proteiner som täcker, skyddar och mobiliserar dem.
I prokaryoter är DNA associerat med proteiner som bestämmer bildandet av komplexa öglor i en struktur som kallas nukleoid. Den maximala nivån av DNA-komprimering i en eukaryot cell är å andra sidan den mitotiska eller meiotiska kromosomen..
Det enda fallet där ett B-DNA inte är förpackat är ett forskningslaboratorium som strävar efter detta syfte..
Artikelindex
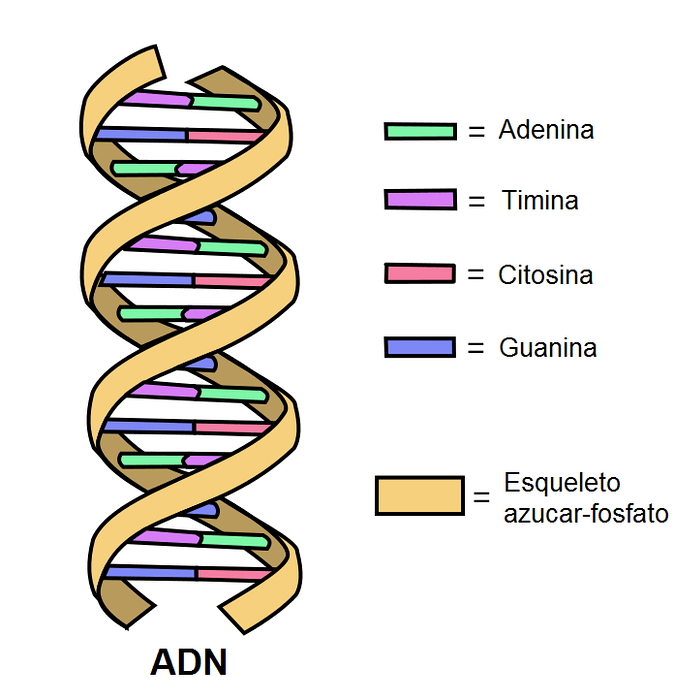
DNA består av två antiparallella band som bildar en dubbel spiral. Var och en av dem har ett fosfodiesterbindningsskelett på vilket socker kopplat till kvävebaser är fästa..
Inuti molekylen bildar kvävebaserna i ett band vätebindningar (två eller tre) med det komplementära bandet.
I en molekyl som denna visar de flesta viktiga bindningsvinklar fri rotation. Kvävebas-socker, socker-fosfatgrupp och fosfodiesterbindningar är flexibla.
Detta gör det möjligt för DNA, sett som en flexibel stav, att visa en viss förmåga att böja och vrida. Denna flexibilitet gör det möjligt för DNA att anta komplexa lokala strukturer och bilda interaktionsöglor på korta, medelstora och långa avstånd..
Denna flexibilitet förklarar också hur 2 meter DNA kan upprätthållas i varje diploid cell hos en människa. I en könsceller (haploida cell) skulle det vara en meter DNA.
Även om det inte är en obrytbar regel, existerar bakteriekromosomen som en enda superlindad dubbelbandig DNA-molekyl..
Den dubbla helixen vrids mer på sig själv (mer än 10 bp per varv) vilket ger en viss komprimering. Lokala knop genereras också tack vare manipulationer som styrs enzymatiskt.
Dessutom finns det sekvenser i DNA som gör att domäner kan bildas i stora öglor. Vi kallar strukturen som härrör från superspolen och ordnade öglor en nukleoid..
Dessa genomgår dynamiska förändringar tack vare vissa proteiner som ger en viss strukturell stabilitet för den komprimerade kromosomen. Graden av komprimering i bakterier och archaea är så effektiv att det kan finnas mer än en kromosom per nukleoid.
Nukleoid komprimerar det prokaryota DNA: t minst 1000 gånger. Den mycket topologiska strukturen av nukleoid är en grundläggande del av regleringen av gener som bärs av kromosomen. Det vill säga struktur och funktion utgör samma enhet..
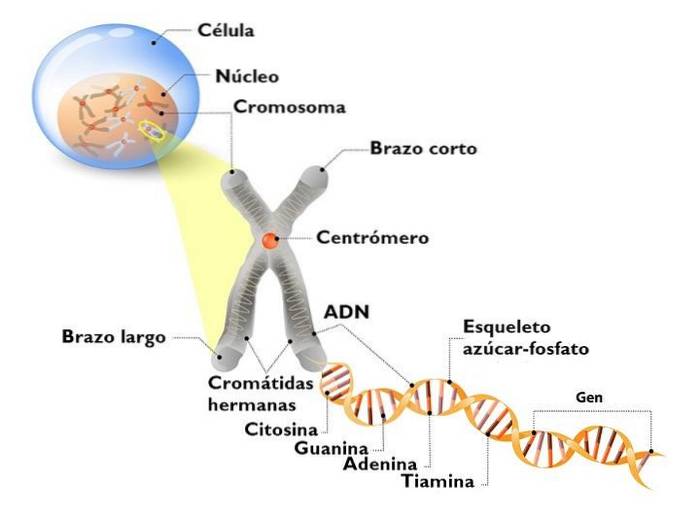
DNA i den eukaryota kärnan är inte naken. Det interagerar med många proteiner, varav det viktigaste är histoner. Histoner är små, positivt laddade proteiner som binder till DNA icke-specifikt.
I kärnan ser vi ett komplext DNA: histoner, som vi kallar kromatin. Det högt kondenserade kromatinet, som vanligtvis inte uttrycks, är heterokromatin. Däremot är det minst komprimerade (lösare), eller eukromatin, kromatin med gener som uttrycks.
Kromatin har olika nivåer av komprimering. Det mest elementära är nukleosomens; Det följs av solenoidfibrerna och interfaskromatinöglorna. Det är först när en kromosom delar sig att maximala komprimeringsnivåer visas..
Nukleosomen är den grundläggande organisationsenheten för kromatin. Varje nukleosom består av en oktamer av histoner som bildar en slags trumma..
Oktamern består av två kopior av var och en av histonerna H2A, H2B, H3 och H4. Runt dem går DNA cirka 1,7 gånger. Det följs av en fraktion av fritt DNA som kallas en 20 bp linker associerad med histon H1 och sedan en annan nukleosom. Mängden DNA i en nukleosom och det som binder den till en annan är cirka 166 baspar.
Detta DNA-förpackningssteg komprimerar molekylen cirka 7 gånger. Det vill säga vi går från en meter till drygt 14 cm DNA.
Denna packning är möjlig eftersom de positiva histonerna avlägsnar den negativa laddningen av DNA och den därav följande elektrostatiska självåterstötningen. Den andra anledningen är att DNA kan vikas på ett sådant sätt att det kan vända oktameren i histoner..
Fibrerna av pärlor i ett halsband som bildas av många successiva nukleosomer lindas ytterligare in i en mer kompakterad struktur.
Även om vi inte är tydliga med vilken struktur den faktiskt antar, vet vi att den når en tjocklek på cirka 30 nm. Detta är den så kallade 30 nm fibern; Histon H1 är väsentlig för dess bildning och stabilitet.
30 nm fibern är den grundläggande strukturella enheten för heterokromatin. Det för laxa nukleosomer, det för eukromatin.
30 nm fibern är dock inte helt linjär. Tvärtom bildar den slingor med en längd på cirka 300 nm på ett serpentint sätt på en lite känd proteinmatris..
Dessa öglor på en proteinmatris bildar en mer kompakt kromatinfiber med en diameter av 250 nm. Slutligen inriktas de som en enda spiral på 700 nm tjock, vilket ger upphov till en av systerkromatiderna i en mitotisk kromosom..
I slutändan komprimeras DNA i kärnkromatin cirka 10 000 gånger på kromosomen i den delande cellen. I mellanfaskärnan är dess komprimering också hög eftersom den är cirka 1000 gånger jämfört med det "linjära" DNA..

I utvecklingsbiologins värld sägs gametogenes återställa epigenomen. Det vill säga, det raderar DNA-märkena att livet för den person som gav upphov till den gamete som producerades eller upplevdes.
Dessa taggar inkluderar DNA-metylering och kovalenta modifieringar av histoner (kod för histoner). Men inte hela epigenomen återställs. Vad som återstår med märken kommer att vara ansvarig för faderns eller moderns genetiska avtryck.
Den implicita återställningen till gametogenes är lättare att se i spermier. I spermier är DNA inte packat med histoner. Därför ärvs inte informationen som är associerad med dess modifieringar i producentorganismen..
I spermier förpackas DNA genom interaktion med icke-specifika DNA-bindande proteiner som kallas protaminer. Dessa proteiner bildar disulfidbindningar med varandra, vilket hjälper till att bilda överlappande DNA-skikt som inte stöter bort elektrostatiskt..



Ingen har kommenterat den här artikeln än.