De replikeringsgaffel Det är den punkt vid vilken DNA-replikation sker, det kallas också en tillväxtpunkt. Den är Y-formad och när replikering sker rör sig hårnålen genom DNA-molekylen.
DNA-replikering är den cellulära processen som involverar duplicering av genetiskt material i cellen. DNA-strukturen är en dubbel helix, och för att replikera dess innehåll måste den öppnas. Var och en av trådarna kommer att vara en del av den nya DNA-kedjan, eftersom replikering är en halvkonservativ process.
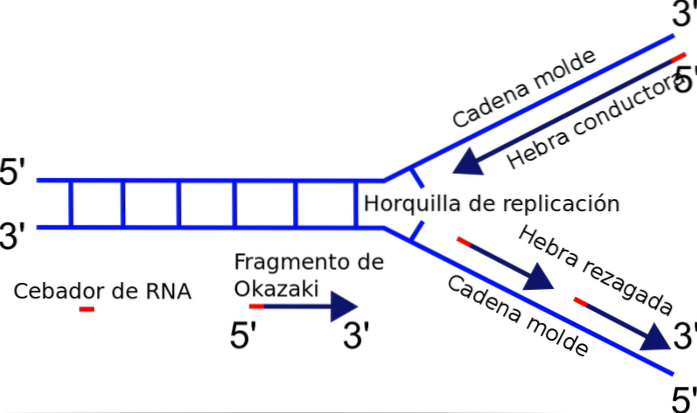
Replikeringsgaffeln bildas exakt mellan korsningen mellan den nyligen separerade mallen eller mallsträngarna och det duplex-DNA som ännu inte har duplicerats. När DNA-replikering initieras kan en av trådarna enkelt dupliceras, medan den andra strängen står inför ett polaritetsproblem..
Enzymet som ansvarar för att polymerisera kedjan - DNA-polymeras - syntetiserar endast DNA-strängen i 5'-3'-riktningen. Således är den ena strängen kontinuerlig och den andra genomgår diskontinuerlig replikering och genererar Okazaki-fragment..
Artikelindex
DNA är molekylen som lagrar nödvändig genetisk information för alla levande organismer - med undantag för vissa virus.
Denna enorma polymer bestående av fyra olika nukleotider (A, T, G och C) finns i kärnan av eukaryoter, i var och en av cellerna som utgör vävnaderna hos dessa varelser (utom i mogna röda blodkroppar hos däggdjur, som saknar kärna).
Varje gång en cell delar sig måste DNA replikeras för att skapa en dottercell med genetiskt material.
Replikering kan vara enkelriktad eller dubbelriktad, beroende på bildandet av replikationsgaffeln vid utgångspunkten..
Logiskt, i fallet med replikering i en riktning bildas endast en hårnål, medan i dubbelriktad replikering bildas två hårnålar.
För denna process är ett komplext enzymatiskt maskineri nödvändigt, som fungerar snabbt och som kan replikera DNA exakt. De viktigaste enzymerna är DNA-polymeras, DNA-primas, DNA-helikas, DNA-ligas och topoisomeras..
DNA-replikering börjar inte på något slumpmässigt ställe i molekylen. Det finns specifika regioner i DNA som markerar början på replikering.
I de flesta bakterier har bakteriekromosomen en enda AT-rik startpunkt. Denna komposition är logisk, eftersom det underlättar öppningen av regionen (AT-paren är förenade med två vätebindningar, medan GC-paret med tre).
När DNA börjar öppna, bildas en Y-formad struktur: replikationsgaffeln..
DNA-polymeras kan inte starta dotterkedjesyntes från grunden. Du behöver en molekyl som har en 3'-ände så att polymeraset har var man ska börja polymerisera.
Denna fria 3'-ände tillhandahålls av en liten nukleotidmolekyl som kallas primer. Den första fungerar som en slags krok för polymeraset.
Under replikationen har replikeringsgaffeln förmågan att röra sig längs DNA: t. Genomgången av replikationsgaffeln lämnar två enkelbandiga DNA-molekyler som styr bildandet av dubbelbandets dottermolekyler..
Hårnålen kan gå vidare tack vare effekten av helikasenzymer som avlindar DNA-molekylen. Detta enzym bryter vätebindningarna mellan baspar och möjliggör hårnålsförskjutning..
Replikeringen är klar när de två hårnålarna är vid 180 ° C från ursprunget.
I det här fallet pratar vi om hur replikationsprocessen flyter i bakterier och det är nödvändigt att markera hela torsionsprocessen för den cirkulära molekylen som replikering innebär. Topoisomeraser spelar en viktig roll i avvecklingen av molekylen.
Har du någonsin undrat hur replikering sker i DNA? Det vill säga en annan dubbelhelix måste komma ut ur dubbelhelixen, men hur händer det? Under flera år var detta en öppen fråga bland biologer. Flera permutationer kan existera: två gamla strängar tillsammans och två nya tillsammans, eller en ny tråd och en gammal tråd för att bilda den dubbla spiralen..
1957 besvarades denna fråga av forskarna Matthew Meselson och Franklin Stahl. Den replikeringsmodell som författarna föreslog var den halvkonservativa.
Meselson och Stahl hävdade att resultatet av replikering är två DNA-dubbla helixmolekyler. Var och en av de resulterande molekylerna består av en gammal sträng (från moder- eller initialmolekylen) och en ny syntetiserad ny sträng.
DNA-helixen består av två kedjor som kör antiparallell: en går i 5'-3'-riktningen och den andra 3'-5 '.
Det mest framträdande enzymet i replikationsprocessen är DNA-polymeras, som är ansvarigt för att katalysera föreningen av de nya nukleotiderna som kommer att läggas till kedjan. DNA-polymeras kan endast förlänga kedjan i 5'-3'-riktningen. Detta faktum hindrar samtidig duplicering av kedjorna i replikationsgaffeln..
Varför? Tillsatsen av nukleotider sker vid den fria änden 3 där en hydroxylgrupp (-OH) finns. Således kan endast en av trådarna lätt amplifieras genom terminal tillsats av nukleotiden till 3'-änden. Detta kallas en ledande eller kontinuerlig sträng.
Den andra strängen kan inte förlängas, eftersom den fria änden är 5 'och inte 3' och varken polymeras katalyserar tillsatsen av nukleotider till 5 '-änden. Problemet löses med syntes av flera korta fragment (130 till 200 nukleotider), var och en i normal replikationsriktning från 5 'till 3'.
Denna diskontinuerliga syntes av fragment slutar med förening av var och en av delarna, en reaktion katalyserad av DNA-ligas. För att hedra upptäckaren av denna mekanism, Reiji Okazaki, kallas de små syntetiserade segmenten Okazaki-fragment..


Ingen har kommenterat den här artikeln än.