A fylogenetiskt träd det är en grafisk matematisk framställning av historien och släktförhållandena mellan grupper, populationer, arter eller någon annan taxonomisk kategori. Teoretiskt sett kan alla fylogenetiska träd grupperas i livets träd och utgör det universella trädet.
Dessa grafiska framställningar har revolutionerat studien av evolutionär biologi, eftersom de möjliggör etablering och definiering av en art, testning av olika evolutionära hypoteser (såsom endosymbiotisk teori), utvärdering av sjukdomars ursprung (såsom HIV) etc..
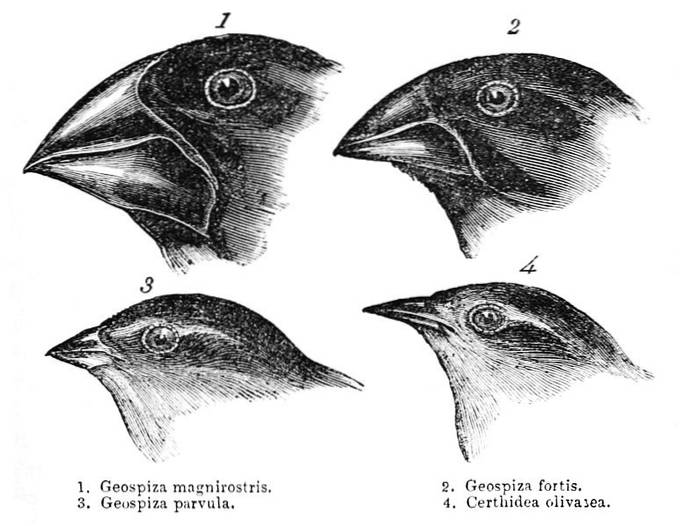
Träd kan rekonstrueras med morfologiska eller molekylära tecken, eller båda. På samma sätt finns det olika metoder för att bygga dem, den vanligaste är den kladistiska metoden. Detta syftar till att identifiera delade härledda karaktärer, så kallade synapomorphy.
Artikelindex
En av de principer som utvecklats av Charles Darwin utgör den gemensamma anor för alla levande organismer - det vill säga vi delar alla en avlägsen förfader.
På "Arternas ursprung" Darwin lyfter metaforen för "livets träd". I själva verket använder han ett hypotetiskt grafiskt träd för att utveckla sin idé (märkligt nog är det den enda bilden av Källa).
Representationen av denna metafor är vad vi känner till fylogenetiska träd, som gör det möjligt för oss att grafiskt visa historien och förhållandena för en viss grupp av organismer..
I fylogenetiska träd kan vi skilja på följande delar - fortsätter med den botaniska analogin:
Grenar: Trädets linjer kallas ”grenar” och dessa representerar studiepopulationerna över tiden. Beroende på typen av träd (se nedan) kan grenens längd kanske inte ha någon betydelse.
På grenarnas tips hittar vi de organismer som vi vill utvärdera. Dessa kan vara enheter som för närvarande lever eller utdöda varelser. Arten skulle vara löv från vårt träd.
Rot: roten är trädets äldsta gren. Vissa har det och kallas rotade träd, medan andra inte gör det.
Knutpunkter: grenarnas grenpunkter i två eller flera släkter kallas noder. Pricken representerar den senaste gemensamma förfadern för efterkommande grupper (notera att dessa förfäder är hypotetiska).
Förekomsten av en nod innebär en specieringshändelse - skapande av nya arter. Efter detta följer varje art sin evolutionära kurs.
Förutom dessa tre grundläggande begrepp finns det andra nödvändiga termer när det gäller fylogenetiska träd:
Politomi: när ett fylogenetiskt träd har mer än två grenar i en nod, sägs det att det finns en polytomi. I dessa fall är det fylogenetiska trädet inte helt löst, eftersom förhållandena mellan de involverade organismerna inte är tydliga. Detta beror vanligtvis på brist på data och kan bara åtgärdas när en forskare samlar mer.
Extern grupp: i fylogenetiska ämnen är det vanligt att höra begreppet extern grupp - även kallad utgrupp. Denna grupp är vald för att kunna rota trädet. Det bör väljas som en taxon som tidigare skilde sig från studiegruppen. Om jag till exempel studerar tagghudar kan du använda utgrupp havet sprutar.
Det finns tre grundläggande typer av träd: kladogram, tillsatssträd och ultrametriska träd..
Kladogram är de enklaste träden och visar organismernas förhållande i termer av gemensamma anor. Informationen om denna typ av träd finns i förgreningsmönstren, eftersom grenarnas storlek inte har någon ytterligare betydelse.
Den andra typen av träd är tillsatsen, även kallad metriska träd eller fylogram. Grenarnas längd är relaterad till mängden evolutionär förändring.
Slutligen har vi ultrametriska träd eller dendogram, där alla trädens spetsar ligger på samma avstånd (vilket inte är fallet i fylogrammet, där en spets kan verka lägre eller högre än sin partner). Grenens längd är relaterad till evolutionstiden.
Valet av träd är direkt relaterat till den evolutionära frågan som vi vill svara på. Till exempel, om vi bara bryr oss om relationerna mellan individer, är ett kladogram tillräckligt för studien.
Även om fylogenetiska träd ofta används i stor utsträckning i evolutionär biologi (och allmän biologi), finns det många studenter och utövare som misstolkar budskapet som dessa till synes enkla grafer är avsedda att leverera till läsaren..
Det första misstaget är att läsa dem i sidled, förutsatt att evolutionen innebär framsteg. Om vi förstår evolutionsprocessen korrekt finns det ingen anledning att tro att de förfäderna är till vänster och de mer avancerade arterna till höger..
Även om den botaniska analogin av ett träd är mycket användbar, kommer det en punkt där det inte längre är så exakt. Det finns en viktig trädstruktur som inte finns i trädet: stammen. I fylogenetiska träd hittar vi inga huvudgrenar.
Specifikt kan vissa människor betrakta människan som det ultimata "målet" för evolutionen, och därför arten. Homo sapiens bör alltid lokaliseras som slutlig enhet.
Denna uppfattning överensstämmer dock inte med evolutionära principer. Om vi förstår att fylogenetiska träd är rörliga element kan vi placera Homo i någon terminal position av trädet, eftersom denna egenskap inte är relevant i representationen.
En viktig egenskap som vi måste förstå om fylogenetiska träd är att de representerar icke-statiska grafer..
I dem kan alla dessa grenar rotera - på samma sätt som en mobil kan göra det. Vi menar inte att vi kan flytta grenarna efter behag, eftersom vissa rörelser skulle innebära förändring av mönstret eller topologi Från trädet. Vad vi kan rotera är noderna.
För att tolka ett träds budskap får vi inte fokusera på grenarnas spetsar, vi måste fokusera på grenpunkterna, som är den viktigaste aspekten av grafen.
Dessutom måste vi komma ihåg att det finns flera sätt att rita ett träd. Många gånger beror det på stilen på boken eller tidningen och förändringar i grenarnas form och placering påverkar inte informationen de vill förmedla till oss..
När ska vi hänvisa till arter nuvarande vi bör inte tillämpa förfäderns konnotationer till dem. Till exempel, när vi tänker på förhållandena mellan schimpanser och människor, kunde vi felaktigt förstå att schimpanser är förfäder med avseende på vår släktlinje..
Men den gemensamma förfadern för schimpanser och människor var ingen av dem. Att tro att schimpansen är förfäder skulle vara att anta att dess utveckling stannade när båda släktarna hade separerat..
Efter samma logik som dessa idéer berättar ett fylogenetiskt träd inte om det finns unga arter heller. Eftersom de allelfrekvenserna är i konstant förändring och det finns nya tecken som förändras över tiden är det svårt att bestämma åldern hos en art och, säkert, ett träd ger oss inte sådan information.
"Förändringen i allelfrekvenser över tiden" är hur populationsgenetik definierar evolution.
När vi tittar på ett fylogenetiskt träd måste vi förstå att denna graf helt enkelt är en hypotes som genereras från konkreta bevis. Det kan vara så att om vi lägger till fler tecken i trädet så ändrar det dess topologi.
Forskarnas expertis när det gäller att välja de bästa karaktärerna för att klargöra förhållandena mellan organismerna i fråga är nyckeln. Dessutom finns det mycket kraftfulla statistiska verktyg som gör det möjligt för forskare att utvärdera träd och välja det mest troliga..
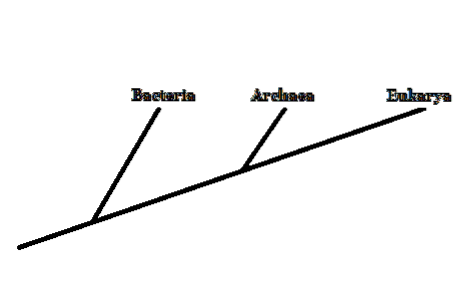
1977 föreslog forskaren Carl Woese att gruppera levande organismer i tre områden: Archaea, Bacteria och Eukarya. Detta nya klassificeringssystem (tidigare fanns det bara två kategorier, Eukaryota och Prokaryota) baserades på den ribosomala RNA-molekylmarkören.
Bakterier och eukaryoter är allmänt kända organismer. Archaea förväxlas ofta med bakterier. Dessa skiljer sig emellertid djupt i strukturen hos deras cellulära komponenter..
Därför, även om de är mikroskopiska organismer som bakterier, är medlemmar av Archaea-domänen närmare besläktade med eukaryoter - eftersom de delar en närmare gemensam förfader..
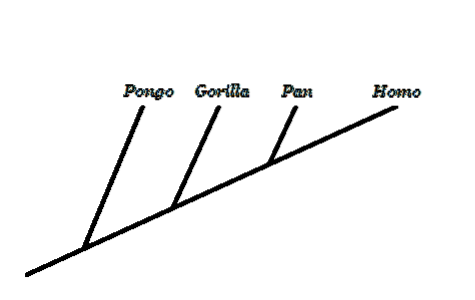
Inom evolutionär biologi är ett av de mest kontroversiella ämnena människans utveckling. För motståndarna till denna teori är en utveckling med utgångspunkt från en apelikal förfader som gav upphov till den moderna människan inte logisk..
Ett nyckelbegrepp är att förstå att vi inte utvecklats från nuvarande apor, utan vi delar en gemensam förfader med dem. I apans och människans träd sticker det ut att det vi känner till "apa" inte är en giltig monofyletisk grupp, eftersom det utesluter människor.
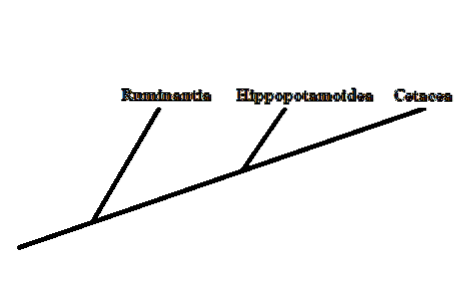
Evolutionärt sett representerade valar en grupp ryggradsdjur vars förhållanden med resten av sina andra däggdjur inte var så tydliga. Morfologiskt har valar, delfiner och andra medlemmar få likheter med resten av däggdjur.
För närvarande har det, tack vare studien av olika morfologiska och molekylära karaktärer, varit möjligt att dra slutsatsen att systergruppen för de stora valarna bildas av artiodaktylerna - hovdjur med jämna hovar..



Ingen har kommenterat den här artikeln än.