De homolog rekombination det är en process som involverar utbytet av DNA-molekyler mellan liknande eller identiska delar av genomet. Celler använder homolog rekombination främst för att reparera pauser i genetiskt material, vilket genererar genetisk variation i populationer.
I allmänhet involverar homolog rekombination den fysiska parningen mellan homologa områden i det genetiska materialet, följt av att kedjorna som kommer att genomgå utbyte bryts och slutligen föreningen av de nya kombinerade DNA-molekylerna..
Brott i DNA måste repareras så snabbt och effektivt som möjligt. När skadan inte repareras kan konsekvenserna bli allvarliga och till och med dödliga. Hos bakterier är den huvudsakliga funktionen för homolog rekombination att reparera dessa avbrott i det genetiska materialet.
Homolog rekombination anses vara en av de viktigaste mekanismerna som möjliggör genomets stabilitet. Det finns i alla livets områden och även i virus, så det är förmodligen en vital mekanism som uppträdde mycket tidigt i livets utveckling.
Artikelindex
En av de mest relevanta principerna som föreslås av Gregor Mendel består av självständighet i segregeringen av karaktärer. Enligt denna lag överförs de olika generna från förälder till barn oberoende.
Men 1900 var det mycket tydliga undantag från denna princip. De engelska genetikerna Bateson och Punnett visade att många gånger ärftliga karaktärer ärvs tillsammans, och för dessa egenskaper har principen som Mendel förkunnat ingen giltighet..
Senare undersökningar lyckades belysa förekomsten av rekombinationsprocessen, där celler kunde utbyta genetiskt material. I de fall där generna ärvde utbyttes inte DNA på grund av den fysiska närheten mellan generna..
Homolog rekombination är ett cellulärt fenomen som involverar det fysiska utbytet av DNA-sekvenser mellan två kromosomer. Rekombination involverar en uppsättning gener som kallas gener rec. Dessa kodar för olika enzymer som deltar i processen.
DNA-molekyler betraktas som "homologa" när de delar liknande eller identiska sekvenser av mer än 100 baspar. DNA har små regioner som kan skilja sig från varandra, och dessa varianter kallas alleler.
I levande saker betraktas allt DNA som rekombinant DNA. Utbytet av genetiskt material mellan kromosomer sker kontinuerligt, blandning och omläggning av generna på kromosomerna.
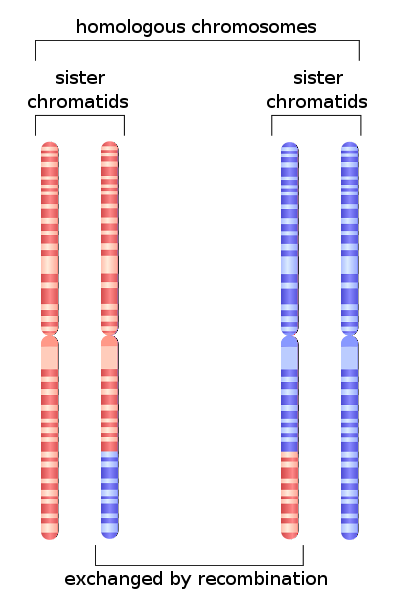
Denna process sker uppenbarligen i meios. Specifikt i den fas där kromosomer parar sig i den första celldelningen. I detta skede sker utbytet av genetiskt material mellan kromosomer.
Historiskt betecknas denna process i litteraturen med det angelsaxiska ordet korsa över. Denna händelse är ett av resultaten av homolog rekombination..
Frekvensen av korsa över mellan två gener av samma kromosom beror främst på avståndet som finns mellan dem; ju mindre det fysiska avståndet mellan dem desto lägre utbytesfrekvens.
Genetiskt material utsätts ständigt för skador orsakade av endogena och exogena källor, såsom strålning, till exempel.
Mänskliga celler uppskattas ha ett betydande antal DNA-lesioner i storleksordningen tio till hundratals per dag. Dessa skador måste repareras för att undvika potentiella skadliga mutationer, replikations- och transkriptionsblock och skador på kromosomnivå..
Ur medicinsk synvinkel resulterar DNA-skador som inte repareras korrekt i utvecklingen av tumörer och andra patologier.
Homolog rekombination är en händelse som möjliggör DNA-reparation, vilket möjliggör återvinning av förlorade sekvenser med den andra (homologa) DNA-strängen som mall..
Denna metaboliska process är närvarande i alla livsformer, vilket ger en högkvalitativ mekanism som möjliggör reparation av "luckor" i DNA, dubbelsträngade brytningar och tvärbindningar mellan DNA-strängar..
En av de mest relevanta konsekvenserna av rekombination är genereringen av ny genetisk variation. Tillsammans med mutationer är de de två processerna som genererar variation hos levande varelser - kom ihåg att variation är råmaterialet för evolutionen.
Dessutom tillhandahåller den en mekanism för att återställa replikeringsgafflar som har skadats..
Hos bakterier förekommer ofta horisontella genöverföringshändelser. Dessa klassificeras som konjugering, transformation och transduktion. Här tar prokaryoter DNA från en annan organism och till och med från olika arter.
Under dessa processer sker homolog rekombination mellan mottagarcellen och givarcellen..
Homolog rekombination börjar med paus i en av strängarna i den kromosomala DNA-molekylen. Efter detta inträffar en serie steg katalyserade av flera enzymer..
3'-änden där skärningen inträffar invaderas av den homologa dubbla DNA-strängen. Invasionsprocessen är avgörande. Med "homolog kedja" menar vi delarna av kromosomerna som har samma gener i en linjär ordning, även om nukleotidsekvenserna inte behöver vara identiska.
Denna invasion av strängen placerar homologa kromosomer mot varandra. Detta fenomen att möta strängar kallas synaps (inte förväxlas med synaps i neuroner, här används termen med en annan betydelse).
Synapsen innebär inte nödvändigtvis en direkt kontakt mellan båda homologa sekvenserna, DNA kan fortsätta att röra sig en stund tills den hittar den homologa delen. Denna sökprocess kallas homolog inriktning..
Sedan inträffar en händelse som kallas "strandinvasion". En kromosom är en dubbel DNA-helix. Vid homolog rekombination letar två kromosomer efter sina homologa sekvenser. I en av spiralerna separeras trådarna och denna tråd "invaderar" den dubbla spiralstrukturen och bildar strukturen som kallas D-slingan..
D-loop-kedjan har förskjutits av invasionen av brytsträngen och parar ihop med den kompletterande strängen i den ursprungliga dubbelspiralen.
Nästa steg är bildandet av Holliday-fackföreningarna. Här är ändarna på de utbytta trådarna bundna ihop. Denna union har förmågan att röra sig i vilken riktning som helst. Leden kan gå sönder och bildas flera gånger.
Den sista processen för rekombination är upplösningen av dessa fackföreningar och det finns två sätt eller sätt på vilka cellen uppnår detta. En av dem är klyvning av unionen eller genom en process som kallas upplösning, typisk för eukaryota organismer..
I den första mekanismen regenererar brytningen av Holliday-korsningen två kedjor. I den andra händelsen "upplösning" inträffar ett slags kollaps i unionen..
Ett viktigt protein i rekombinationsprocessen kallas Rad51 i eukaryota celler och RecA in Escherichia coli. Det fungerar i de olika faserna av rekombination: före, under och efter synaps.
Rad51-proteinet underlättar bildandet av den fysiska kopplingen mellan det invaderande DNA: t och det tempererade DNA: t. I denna process genereras heteroduplex-DNA.
Rad51, och dess RecA-motsvarighet, katalyserar sökandet efter homologt DNA och utbytet av DNA-strängar. Dessa proteiner har förmågan att samarbetsvilligt binda till enkelband-DNA..
Det finns också paraloggener (som härrör från gendupliceringshändelser i en organisation) av Rad51, kallad Rad55 och Rad57. Hos människor har fem Rad51-paraloggener identifierats som kallas Rad51B, Rad51C, Rad51D, Xrcc2 och Xrcc3..
Eftersom rekombination kräver fysisk bindning på kromosomer är det ett viktigt steg i korrekt segregering under meios. Om korrekt rekombination inte sker kan resultatet bli stor patologi.
Icke-separering av kromosomer eller fel i segregering är en av de vanligaste orsakerna till aborter och anomalier av kromosomalt ursprung, såsom trisomi av kromosom 21, vilket orsakar Downs syndrom..
Även om rekombination vanligtvis är en ganska exakt process är regioner i genomet som upprepas och gener som har flera kopior genom genomet element som är benägna att ojämn delning.
Denna avel producerar olika kliniskt relevanta egenskaper, inklusive vanliga sjukdomar som talassemi och autism.
Molekylära biologer har utnyttjat kunskapen om mekanismen för homolog rekombination för att skapa olika tekniker. En av dessa möjliggör skapandet av organismer "knockout-".
Dessa genetiskt modifierade organismer gör det möjligt att klargöra funktionen av en gen av intresse.
En av metoderna som används för att skapa knockouts Den består av undertryckande av uttrycket för den specifika genen och ersätter den ursprungliga genen med en modifierad eller "skadad" version. Genen byts mot den muterade versionen genom homolog rekombination.
Förutom homolog eller legitim rekombination finns det andra typer av utbyte av genetiskt material.
När de regioner av DNA som byter material är icke-alleliska (homologa kromosomer) är resultatet duplicering eller reduktion av gener. Denna process är känd som icke-homolog rekombination eller ojämn rekombination..
Tillsammans kan genetiskt material också utbytas mellan systerkromatider på samma kromosom. Denna process inträffar både i den meiotiska och mitotiska uppdelningen och kallas ojämnt utbyte..



Ingen har kommenterat den här artikeln än.