De Cyklisk AMP eller adenosin 3 ', 5'-monofosfat är en cyklisk nukleotid som fungerar som en sekundär budbärare och är en del av de grundläggande elementen i biokemisk kontroll och intracellulär kommunikation i många levande organismer.
Dess existens demonstrerades för nästan 70 år sedan av Sutherland och Rall (1958), som beskrev fenomenet ackumulering av denna nukleotid i leverceller som ett resultat av administrering av adrenalin (adrenalin).
Sedan dess upptäckt har cyklisk AMP associerats i däggdjur med effekten av många hormoner, med endokrin och exokrin utsöndring, med frisättning av neurotransmittorer vid synaps och neuromuskulära korsningar, bland många andra funktioner..
Dess syntes katalyseras av tre familjer av proteiner associerade med plasmamembranet känt som adenylcyklas eller adenylatcyklas, som kan producera den cykliska föreningen från ATP och frigöra pyrofosfat i cellen..
Å andra sidan medieras dess nedbrytning av enzymer från fosfodiesterasfamiljen, vilka är lösliga proteiner som huvudsakligen finns i cytosolen..
Dessa enzymer, och därför cykliska AMP, finns i extremt olika organismer, så enkla som encelliga alger och många andra mikroorganismer (bakterier och andra) och lika komplexa som multicellulära djur med komplicerade signalvägar..
Även om dess närvaro i växter är en fråga om diskussion, finns det vissa bevis som indikerar att vissa växtarter har adenylatcyklasaktivitet, även om dess funktion inte har fastställts tillfredsställande..
Artikelindex
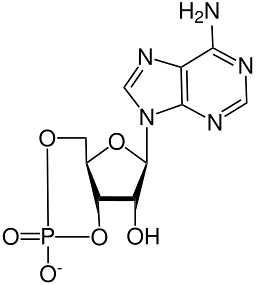
Den kemiska strukturen för cyklisk AMP har belysts genom röntgenkristallografi och genom protonkärnmagnetiska resonansstudier..
Detta är en värmestabil cyklisk molekyl ("värmestabil ") och mer stabil mot alkalisk hydrolys än dess icke-cykliska motsvarighet, AMP eller adenosinmonofosfat.
Liksom alla fosfatnukleotider har cyklisk AMP en fosfatgrupp bunden till kolsyre vid 5'-positionen hos en ribosmolekyl, som i sin tur är bunden till en heterocyklisk kvävebas via kolet i 1'-positionen och det motsvarar en adenin.
Fosfatgruppen i ribossocker, till skillnad från de icke-cykliska fosfatnukleotiderna, smälter samman vid trans genom en fosfodiesterbindning med oxigenerna från kolerna vid 3 'och 5' positionerna för ribos (3 ', 5'-trans-smält fosfat).
Denna koppling begränsar rörelsen av furanringen som bildar ribos och omsluter fosfatgruppen i en "stol" -konformation..
Jämfört med icke-cykliska nukleotider är cykliska AMP och andra relaterade nukleotider mindre molekyler med lägre polaritet, vilket är en viktig faktor för deras differentiering av proteinerna som svarar på dem..
Konformationen av glykosidbindningen mellan ribos och adeninringen har viss rotationsfrihet. Detta är också en viktig strukturell parameter för dess skillnad från andra nukleotider (inte bara är kvävebasens identitet).
Som en sekundär budbärare deltar cyklisk AMP i aktiveringen av många signalprocesser (efter dess syntes) eller i aktiveringen av olika enzymer "nedströms" i signalkaskaden genom vilken den produceras..
Deltar i glykogenolys i levern och i frisättningen av insulin från bukspottkörteln, i frisättningen av amylas från spottkörtlarna och i effekten av östrogen i livmodern.
Den har universella funktioner i kontrollen av genuttryck och i integrationen av flera metaboliska funktioner. Många cytokiner använder både kalcium och cyklisk AMP för att utföra sina funktioner..
Bland hormonerna som använder cyklisk AMP i sina signalprocesser (antingen genom att öka eller minska dess intracellulära koncentration) kan inkluderas katekolaminer, glukagon, vasopressin, paratyroidhormon, prostaglandiner, insulin, melatonin och adrenalin, bland andra.
En annan av dess många funktioner är att hämma tillväxt, differentiering och proliferation av T-celler i däggdjur, troligen genom aktivering eller induktion av en repressor av de regulatoriska cytokinerna för dessa processer i sådana celler..
Cyklisk AMP och adenylatcyklas som producerar det är också relaterade till funktionen hos många G-proteinkopplade proteinreceptorer, som är associerade med olika signalmekanismer och andra viktiga cellulära processer..
Hos däggdjur har cyklisk AMP funktioner i regleringen av den glykolytiska och glukoneogena vägen genom hämning av aktiviteten hos enzymet fosfofruktokinas 2 (PFK-2), vilket katalyserar den andra reaktionen av glykolys..
Mekanismen involverar deltagande av hormonet glukagon i aktiveringen av hepatisk adenylatcyklas, vilket orsakar en avsevärd ökning av koncentrationen av cykliskt AMP..
Denna cykliska AMP aktiverar ett cAMP-beroende proteinkinas som fosforylerar och inhiberar fosfofruktokinasaktiviteten hos PFK-2, vilket är ett bifunktionellt enzym med fruktosbisfosfatasaktivitet..
En första budbärare (av varierande kemisk natur) som når en specifik cell som en extern stimulans interagerar med ett adenylatcyklasenzym i plasmamembranet, vilket inducerar produktion av cyklisk AMP.
Ökningen av koncentrationen av cykliska AMP-funktioner vid aktivering av andra faktorer (generellt enzymatiska) som har ytterligare funktioner i bland annat inhibering eller aktivering av metaboliska processer eller gentranskription..
En av huvudfunktionerna associerade med denna reglerande molekyl är aktivering av fosforylas- eller kinaseenzymer, som katalyserar tillsatsen eller avlägsnandet av fosforylgrupper till andra proteiner och enzymer..
Normalt åtföljs excitering av en cell av en ökning av koncentrationen av cyklisk AMP, samtidigt med en ökning av transporten av kalcium mot cellen som har funktioner för aktivering av de cykliska AMP-producerande adenylcyklasenzymerna..
Både syntesen och överföringen av meddelandet och nedbrytningen av cyklisk AMP i celler är finreglerade processer som deltar i underhållet av kroppshomeostas.



Ingen har kommenterat den här artikeln än.