De RNA eller RNA (ribonukleinsyra) är en typ av nukleinsyra som finns i eukaryota och prokaryota organismer och i virus. Det är en nukleotidpolymer som innehåller fyra typer av kvävebaser i sin struktur: adenin, guanin, cytosin och uracil.
RNA finns vanligtvis som ett enda band (utom i vissa virus), på ett linjärt sätt eller i en serie komplexa strukturer. Faktum är att RNA har en strukturell dynamik som inte observeras i DNA-dubbelspiralen. De olika typerna av RNA har mycket olika funktioner.
Ribosomala RNA är en del av ribosomer, strukturerna som ansvarar för proteinsyntes i celler. Messenger-RNA fungerar som mellanhänder och transporterar genetisk information till ribosomen, vilket översätter meddelandet från en nukleotidsekvens till en aminosyrasekvens..
Överförings-RNA ansvarar för att aktivera och överföra de olika typerna av aminosyror -20 totalt - till ribosomerna. Det finns en överförings-RNA-molekyl för varje aminosyra som känner igen sekvensen i budbärar-RNA.
Dessutom finns det andra typer av RNA som inte är direkt involverade i proteinsyntes och deltar i genreglering..
Artikelindex
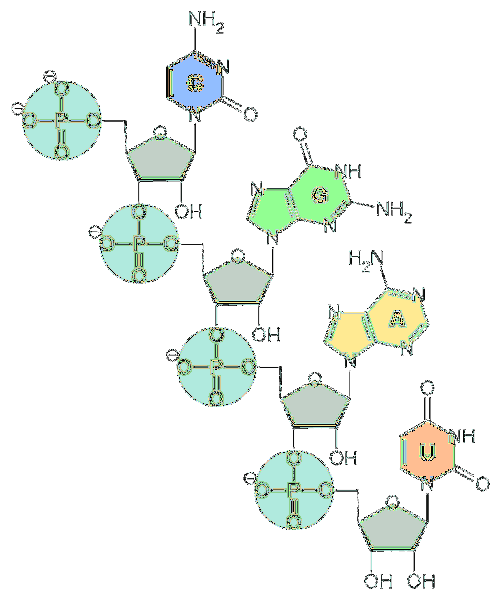
De grundläggande enheterna för RNA är nukleotider. Varje nukleotid består av en kvävehaltig bas (adenin, guanin, cytosin och uracil), en pentos och en fosfatgrupp..
Kvävebaser härrör från två grundläggande föreningar: pyrimidiner och puriner..
Baserna härledda från puriner är adenin och guanin och baserna härrörande från pyrimidiner är cytosin och uracil. Även om dessa är de vanligaste baserna kan nukleinsyror också ha andra typer av baser som är mindre vanliga..
När det gäller pentos är de enheter av d-ribos. Av denna anledning kallas nukleotiderna som utgör RNA "ribonukleotider".
Nukleotider är sammanlänkade genom kemiska bindningar som involverar fosfatgruppen. För att bilda dem är fosfatgruppen vid 5'-änden av en nukleotid bunden till hydroxylgruppen (-OH) vid 3'-änden av nästa nukleotid, vilket skapar en fosfodiesterliknande bindning..
Längs nukleinsyrakedjan har fosfodiesterbindningarna samma orientering. Därför finns det en polaritet hos strängen, som skiljer mellan 3'- och 5'-änden.
Enligt konvention representeras nukleinsyrans struktur av 5'-änden till vänster och 3'-änden till höger.
RNA-produkten från DNA-transkription är ett enkelsträngat band som vänder sig åt höger, i en spiralformad form genom stapling av baser. Interaktionen mellan puriner är mycket större än interaktionen mellan två pyrimidiner på grund av deras storlek.
I RNA är det inte möjligt att tala om en traditionell och referens sekundär struktur, såsom DNA-dubbelspiralen. Den tredimensionella strukturen för varje RNA-molekyl är unik och komplex, jämförbar med den för proteiner (logiskt sett kan vi inte globalisera proteinstrukturen).
Det finns svaga interaktioner som bidrar till stabilisering av RNA, särskilt basstapling, där ringarna är placerade ovanpå varandra. Detta fenomen bidrar också till stabiliteten hos DNA-spiralen..
Om RNA-molekylen hittar en komplementär sekvens kommer de att kunna kopplas ihop och bilda en dubbelsträngad struktur som vänder åt höger. Den dominerande formen är typ A; När det gäller Z-formerna har de bara visats i laboratoriet, medan B-formen inte har observerats.
Generellt finns det korta sekvenser (såsom UUGG) som är belägna i slutet av RNA och som har särheten att bilda öglor stabil. Denna sekvens deltar i vikningen av den tredimensionella strukturen av RNA.
Dessutom kan vätebindningar bildas vid andra ställen än de typiska basparningarna (AU och CG). En av dessa interaktioner sker mellan ribosens 2'-OH och andra grupper.
Att belysa de olika strukturerna som finns i RNA har tjänat till att demonstrera de flera funktionerna hos denna nukleinsyra..
Det finns två klasser av RNA: informativ och funktionell. Den första gruppen inkluderar RNA som deltar i proteinsyntes och fungerar som mellanhänder i processen; informations-RNA är budbärar-RNA.
Däremot ger RNA som tillhör den andra klassen, de funktionella, inte upphov till en ny proteinmolekyl och själva RNA är slutprodukten. Dessa är överförings-RNA och ribosomala RNA..
I däggdjursceller är 80% av RNA ribosomalt RNA, 15% är överförings-RNA och endast en liten del motsvarar budbärar-RNA. Dessa tre typer arbetar tillsammans för att uppnå proteinbiosyntes..
Det finns också små nukleära RNA, små cytoplasmiska RNA och mikroRNA, bland andra. Var och en av de viktigaste typerna kommer att beskrivas i detalj nedan:
I eukaryoter är DNA begränsat till kärnan, medan proteinsyntes sker i cellens cytoplasma, där ribosomer finns. På grund av denna rumsliga separation måste det finnas en medlare som för budskapet från kärnan till cytoplasman och att molekylen är budbärar-RNA..
Messenger RNA, förkortat mRNA, är en mellanliggande molekyl som innehåller information kodad i DNA och som specificerar en aminosyrasekvens som kommer att ge upphov till ett funktionellt protein..
Termen budbärar-RNA föreslogs 1961 av François Jacob och Jacques Monod för att beskriva den del av RNA som överför meddelandet från DNA till ribosomer..
Processen för syntes av ett mRNA från DNA-strängen är känd som transkription och den sker differentiellt mellan prokaryoter och eukaryoter..
Genuttryck styrs av flera faktorer och beror på varje cells behov. Transkription är uppdelad i tre steg: initiering, förlängning och avslutning..
DNA-replikationsprocessen, som sker i varje celldelning, kopierar hela kromosomen. Transkriptionsprocessen är dock mycket mer selektiv, den behandlar endast bearbetning av specifika segment av DNA-strängen och kräver ingen primer..
På Escherichia coli -den bäst studerade bakterien inom biovetenskap - transkription börjar med avvecklingen av DNA-dubbelspiralen och transkriptionsöglan bildas. Enzymet RNA-polymeras är ansvarigt för att syntetisera RNA och när transkriptionen fortsätter återgår DNA-strängen till sin ursprungliga form.
Transkription initieras inte vid slumpmässiga platser på DNA-molekylen; det finns specialiserade webbplatser för detta fenomen, som kallas promotorer. På E coli RNA-polymeras är kopplat några baspar ovanför målområdet.
Sekvenserna där transkriptionsfaktorer är kopplade är ganska konserverade mellan olika arter. En av de mest kända promotorsekvenserna är TATA-rutan.
Vid förlängning adderar RNA-polymerasenzymet nya nukleotider till 3'-OH-änden, efter 5 'till 3' -riktningen. Hydroxylgruppen fungerar som en nukleofil och attackerar alfosfatet i nukleotiden som ska tillsättas. Denna reaktion frigör ett pyrofosfat.
Endast en av DNA-strängarna används för att syntetisera budbärar-RNA: t, som kopieras i 3 'till 5' -riktningen (den antiparallella formen av den nya RNA-strängen). Nukleotiden som ska tillsättas måste överensstämma med baspar: U-par med A och G med C.
RNA-polymeras stoppar processen när den hittar regioner som är rika på cytosin och guanin. Så småningom separeras den nya budbärar-RNA-molekylen från komplexet.
I prokaryoter kan en budbärar-RNA-molekyl koda för mer än ett protein.
När ett mRNA kodar uteslutande för ett protein eller en polypeptid, kallas det monocistronic mRNA, men om det kodar för mer än en proteinprodukt är mRNA polycistronic (notera att i detta sammanhang hänvisar termen cistron till genen).
I eukaryota organismer är den stora majoriteten av mRNA monocistronic och transkriptionsmaskineriet är mycket mer komplext i denna härstamning av organismer. De kännetecknas av att de har tre RNA-polymeraser, betecknade I, II och III, var och en med specifika funktioner.
I ansvarar för att syntetisera pre-rRNA, II syntetiserar budbärar-RNA och några speciella RNA. Slutligen är III ansvarig för överförings-RNA, ribosomal 5S och andra små RNA..
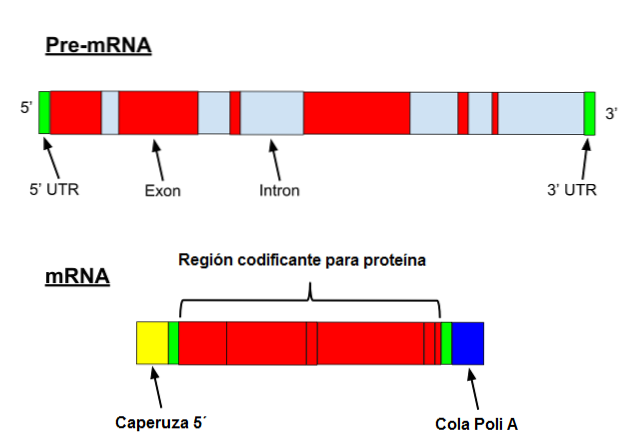
Messenger RNA genomgår en serie specifika modifieringar i eukaryoter. Den första innebär att man lägger till ett "lock" i 5'-änden. Kemiskt är locket en 7-metylguanosinrest bunden till änden med en 5 ', 5'-trifosfatbindning..
Funktionen för denna zon är att skydda RNA från möjlig nedbrytning genom ribonukleaser (enzymer som bryter ner RNA i mindre komponenter).
Dessutom sker avlägsnande av 3'-änden och 80 till 250 adeninrester tillsätts. Denna struktur är känd som polyA "svans" och fungerar som ett bindningsställe för olika proteiner. När en prokaryot förvärvar en polyA-svans tenderar den att stimulera dess nedbrytning..
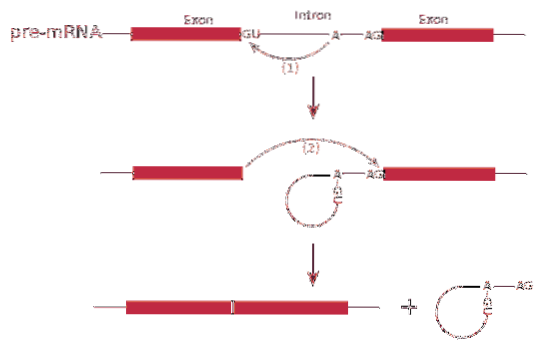
Å andra sidan transkriberas denna budbärare med introner. Introner är DNA-sekvenser som inte är en del av genen men som "avbryter" den sekvensen. Introns översätts inte och måste därför tas bort från budbäraren.
De flesta ryggradsdjurgener har introner, med undantag för gener som kodar för histoner. På samma sätt kan antalet introner i en gen variera från några till tiotals av dessa..
Splicing RNA eller splitsningsprocess involverar avlägsnande av introner i messenger-RNA.
Vissa introner som finns i nukleära eller mitokondriella gener kan utföra processen med skarvning utan hjälp av enzymer eller ATP. Istället utförs processen genom transförestringsreaktioner. Denna mekanism upptäcktes i ciliated protozoan Tetrahymena thermophila.
Däremot finns det en annan grupp budbärare som inte kan förmedla sina egna skarvning, så de behöver ytterligare maskiner. Ett ganska stort antal kärngener tillhör denna grupp..
Processen skarvning det förmedlas av ett proteinkomplex som kallas spliceosom eller splitsningskomplex. Systemet består av specialiserade RNA-komplex som kallas små nukleära ribonukleoproteiner (RNP)..
Det finns fem typer av RNP: U1, U2, U4, U5 och U6, som finns i kärnan och förmedlar processen för skarvning.
De skarvning kan producera mer än en typ av protein - detta kallas skarvning alternativt, eftersom exonerna är ordnade differentiellt, vilket skapar sorter av budbärar-RNA.
Ribosomalt RNA, förkortat rRNA, finns i ribosomer och deltar i proteinbiosyntes. Därför är det en väsentlig komponent i alla celler.
Ribosomalt RNA associeras med proteinmolekyler (cirka 100) för att ge upphov till ribosomala förenheter. De klassificeras beroende på deras sedimenteringskoefficient, betecknad med bokstaven S för Svedberg-enheter..
En ribosom består av två delar: den stora underenheten och den mindre underenheten. Båda subenheterna skiljer sig mellan prokaryoter och eukaryoter när det gäller sedimentationskoefficienten..
Prokaryoter har en stor 50S-underenhet och en liten 30S-underenhet, medan i eukaryoter är den stora underenheten 60S och den lilla 40S.
Generna som kodar för ribosomala RNA finns i kärnan, ett särskilt område i kärnan som inte är avgränsat av ett membran. Ribosomala RNA transkriberas i denna region av RNA-polymeras I.
I celler som syntetiserar stora mängder proteiner; kärnan är en framträdande struktur. Men när cellen i fråga inte kräver ett stort antal proteinprodukter är kärnan en nästan omärkbar struktur.
Den 60S stora ribosomala subenheten är associerad med 28S och 5.8S fragmenten. När det gäller den lilla underenheten (40S) är den associerad med 18S.
I högre eukaryoter kodas pre-rRNA i en 45S-transkriptionsenhet, som involverar RNA-polymeras I. Detta transkript bearbetas till mogna 28S, 18S och 5.8S ribosomala RNA..
När syntesen fortsätter associeras pre-rRNA med olika proteiner och bildar ribonukleoproteinpartiklar. Den genomgår en serie efterföljande modifieringar som inkluderar metylering av 2'-OH-gruppen av ribos och omvandlingen av uridinrester till pseudouridin..
Regionen där dessa förändringar kommer att kontrolleras av mer än 150 små nukleolära RNA-molekyler, som har förmågan att kopplas till pre-rRNA..
I motsats till resten av pre-rRNA transkriberas 5S av RNA-polymeras III i nukleoplasman och inte inuti kärnan. Efter att ha syntetiserats tas det till kärnan för att monteras med 28S och 5.8S, vilket bildar de ribosomala enheterna.
I slutet av monteringsprocessen överförs underenheterna till cytoplasman genom kärnporerna..
Det kan hända att en molekyl av budbärar-RNA ger upphov till flera proteiner samtidigt som binder till mer än en ribosom. När översättningsprocessen fortskrider blir budbärarens ände gratis och kan tas upp av en annan ribosom och startar en ny syntes..
Av denna anledning är det vanligt att hitta ribosomer grupperade (mellan 3 och 10) i en enda molekyl av budbärar-RNA, och denna grupp kallas polyribosom.
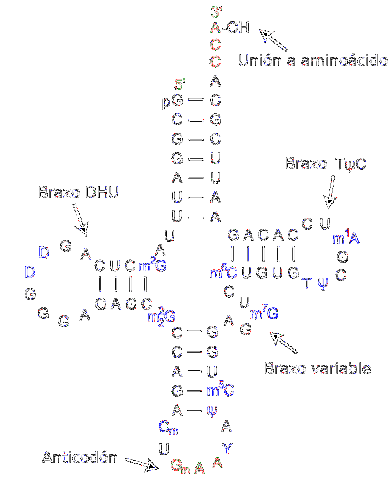
Transfer RNA är ansvarig för överföring av aminosyror när proteinsyntesprocessen fortskrider. De består av cirka 80 nukleotider (jämfört med budbärar-RNA är det en "liten" molekyl).
Strukturen har veck och kors som påminner om en trearmad klöver. En adenylisk ring är belägen vid en av ändarna, där hydroxylgruppen i ribos förmedlar föreningen med aminosyran som ska transporteras.
De olika överförings-RNA: n kombineras uteslutande med en av de tjugo aminosyrorna som utgör proteiner; det är med andra ord fordonet som bär proteinernas byggstenar. Transfer-RNA-komplexet tillsammans med aminosyran kallas aminoacyl-tRNA.
Vidare, i översättningsprocessen - som sker tack vare ribosomer - känner igen varje överförings-RNA ett specifikt kodon i budbärar-RNA. När den känns igen frigörs motsvarande aminosyra och blir en del av den syntetiserade peptiden.
För att känna igen vilken typ av aminosyra som måste levereras har RNA ett "antikodon" beläget i mitten av molekylen. Detta antikodon kan bilda vätebindningar med de komplementära baserna som finns i budbärar-DNA..
MicroRNA eller miRNA är en typ av kort, enkelsträngat RNA, mellan 21 och 23 nukleotider, vars funktion är att reglera expressionen av gener. Eftersom det inte översätts till protein kallas det ofta icke-kodande RNA..
Liksom andra typer av RNA är mikroRNA-bearbetning komplex och involverar ett antal proteiner.
MicroRNA härrör från längre föregångare som kallas mi-pri-RNA, härledda från genens första transkript. I cellens kärna modifieras dessa föregångare i mikroprocessorkomplexet och resultatet är ett pre-miRNA..
Pre-miRNA är 70 nukleotidhårnålar som fortsätter sin bearbetning i cytoplasman av ett enzym som heter Dicer, som monterar det RNA-inducerade tystnadskomplexet (RISC) och slutligen syntetiseras miRNA..
Dessa RNA kan reglera expressionen av gener, eftersom de är komplementära till specifika budbärar-RNA. Genom att para ihop sig med sitt mål kan miRNA: n undertrycka budbäraren eller till och med bryta ner den. Följaktligen kan ribosomen inte översätta nämnda transkript..
En särskild typ av mikroRNA är små interfererande RNA (siRNA), även kallade tysta RNA. De är korta RNA, mellan 20 och 25 nukleotider, vilket hindrar uttrycket av vissa gener.
De är mycket lovande instrument för forskning, eftersom de gör det möjligt att tysta en intressant gen och därmed studera dess möjliga funktion..
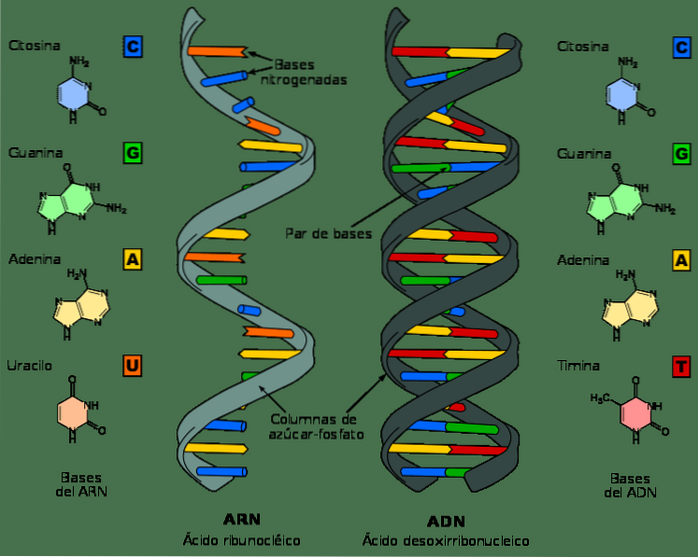
Även om DNA och RNA är nukleinsyror och kan se mycket lika ut vid första anblicken, skiljer de sig åt i flera av sina kemiska och strukturella egenskaper. DNA är en dubbelbandsmolekyl, medan RNA är ettband.
Som ett resultat är RNA en mer mångsidig molekyl och kan anta en mängd olika tredimensionella former. Vissa virus har dock dubbelband-RNA i sitt genetiska material..
I RNA-nukleotider är sockermolekylen en ribos, medan den i DNA är en deoxiribos, som endast skiljer sig i närvaro av en syreatom.
Fosfodiesterbindningen i ryggraden i DNA och RNA är benägen att genomgå en långsam hydrolysprocess utan närvaro av enzymer. Under alkaliska förhållanden hydrolyserar RNA snabbt - tack vare den extra hydroxylgruppen - medan DNA inte gör det.
På liknande sätt är de kvävebaser som utgör nukleotider i DNA guanin, adenin, tymin och cytosin; däremot ersätts tymin i RNA med uracil. Uracil kan paras med adenin, på samma sätt som tymin i DNA.
RNA är den enda kända molekylen som kan lagra information och katalysera kemiska reaktioner samtidigt; Av denna anledning föreslår flera författare att RNA-molekylen var avgörande för livets ursprung. Överraskande nog är ribosomsubstrat andra RNA-molekyler.
Upptäckten av ribozymer ledde till den biokemiska omdefinieringen av "enzym" - sedan tidigare användes termen uteslutande för proteiner med katalytisk aktivitet- och hjälpte till att stödja ett scenario där de första livsformerna endast använde RNA som genetiskt material.


Ingen har kommenterat den här artikeln än.