De kvävehaltiga baser de är organiska föreningar av heterocyklisk form, rika på kväve. De är en del av byggstenarna för nukleinsyror och andra molekyler av biologiskt intresse, såsom nukleosider, dinukleotider och intracellulära budbärare. Med andra ord är kvävebaser en del av enheterna som utgör nukleinsyror (RNA och DNA) och de andra nämnda molekylerna..
Det finns två huvudgrupper av kvävebaser: purin- eller purinbaser och pyrimidin- eller pyrimidinbaser. Adenin och guanin tillhör den första gruppen, medan tymin, cytosin och uracil är pyrimidinbaser. Generellt betecknas dessa baser med deras första bokstav: A, G, T, C och U.
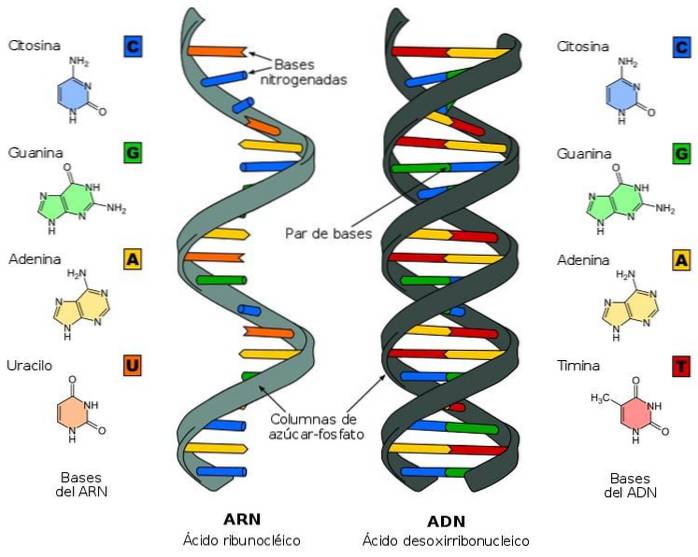
Byggstenarna för DNA är A, G, T och C. I denna ordning av baser kodas all information som krävs för konstruktion och utveckling av en levande organism. I RNA är komponenterna desamma, endast T ersätts med U.
Artikelindex
Kvävebaser är platta molekyler, av aromatisk och heterocyklisk typ, som i allmänhet härrör från puriner eller pyrimidiner..
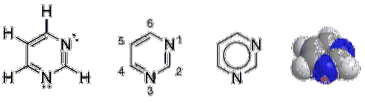
Pyrimidinringen är sexledade heterocykliska aromatiska ringar med två kväveatomer. Atomerna är numrerade medurs.
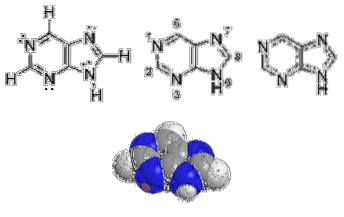
Purinringen består av ett två-ringssystem: en liknar strukturellt pyrimidinringen och den andra liknar imidazolringen. Dessa nio atomer smälts samman i en enda ring.
Pyrimidinringen är ett plant system, medan purinerna avviker något från detta mönster. En liten veck eller rynka har rapporterats mellan imidazolringen och pyrimidinringen..
Inom organisk kemi, a aromatisk ring den definieras som en molekyl vars elektroner från dubbelbindningarna har fri cirkulation inom den cykliska strukturen. Elektronernas rörlighet i ringen ger molekylen stabilitet - om vi jämför den med samma molekyl - men med elektronerna fixerade i dubbelbindningarna..
Den aromatiska karaktären hos detta ringsystem ger dem förmågan att uppleva ett fenomen som kallas keto-enol tautomerism..
Det vill säga puriner och pyrimidiner finns i tautomera par. Keto-tautomerer är dominerande vid neutralt pH för baserna uracil, tymin och guanin. Däremot är enolformen dominerande för cytosin, vid neutralt pH. Denna aspekt är väsentlig för bildandet av vätebindningar mellan baserna..
En annan egenskap hos puriner och pyrimidiner är deras förmåga att starkt absorbera ultraviolett ljus (UV-ljus). Detta absorptionsmönster är en direkt följd av aromatisiteten hos dess heterocykliska ringar..
Absorptionsspektrumet har maximalt nära 260 nm. Forskare använder denna standard för att kvantifiera mängden DNA i sina prover.
Tack vare kvävebasernas starka aromatiska karaktär är dessa molekyler praktiskt taget olösliga i vatten.
Även om det finns ett stort antal kvävebaser, hittar vi bara ett fåtal naturligt i de levande organismernas cellulära miljöer..
De vanligaste pyrimidinerna är cytosin, uracil och tymin (5-metyluracil). Cytosin och tymin är de pyrimidiner som vanligtvis finns i DNA-dubbelspiralen, medan cytosin och uracil är vanliga i RNA. Observera att den enda skillnaden mellan uracil och tymin är en metylgrupp vid kol 5.
På liknande sätt är de vanligaste purinerna adenin (6-aminopurin) och guanin (2-amino-6-oxipurin). Dessa föreningar finns rikligt i både DNA- och RNA-molekylerna..
Det finns andra derivat av puriner som vi hittar naturligt i cellen, inklusive xantin, hypoxantin och urinsyra. De två första kan hittas i nukleinsyror, men på ett mycket sällsynt och specifikt sätt. Däremot finns urinsyra aldrig som en strukturell komponent i dessa biomolekyler..
DNA-strukturen klargjordes av forskarna Watson och Crick. Tack vare deras studie var det möjligt att dra slutsatsen att DNA är en dubbel helix. Den består av en lång kedja av nukleotider kopplade av fosfodiesterbindningar, i vilka fosfatgruppen bildar en brygga mellan hydroxylgrupperna (-OH) i sockerresterna.
Strukturen som vi just har beskrivit liknar en stege tillsammans med respektive ledstång. De kvävehaltiga baserna är analogerna till trappan, som är grupperade i den dubbla spiralen med hjälp av vätebindningar.
I en vätgasbro delar två elektronegativa atomer ett proton mellan baserna. För bildandet av en vätgasbro är det nödvändigt att delta i en väteatom med en liten positiv laddning och en acceptor med en liten negativ laddning..
Bryggan bildas mellan en H och en O. Dessa bindningar är svaga, och de måste vara, eftersom DNA måste öppnas lätt för att replikera.
Baspar bildar vätebindningar enligt följande purin-pyrimidin-parningsmönster känt som Chargaffs regel: guaninpar med cytosin och adeninpar med tymin.
GC-paret bildar tre vätgasbehållare till varandra, medan AT-paret endast är kopplat av två broar. Således kan vi förutsäga att ett DNA med högre GC-innehåll kommer att vara mer stabilt..
Var och en av kedjorna (eller ledstängerna i vår analogi) löper i motsatta riktningar: en 5 '→ 3' och den andra 3 '→ 5'.
Organiska varelser presenterar en typ av biomolekyler som kallas nukleinsyror. Dessa är stora polymerer som består av upprepande monomerer - nukleotider, förenade med en speciell typ av bindning, kallad en fosfodiesterbindning. De klassificeras i två bastyper, DNA och RNA.
Varje nukleotid består av en fosfatgrupp, ett socker (deoxiribostyp i DNA och ribos i RNA) och en av de fem kvävebaserna: A, T, G, C och U. När fosfatgruppen inte är närvarande är molekyl kallas en nukleosid.
DNA är det genetiska materialet hos levande varelser (med undantag för vissa virus som huvudsakligen använder RNA). Med hjälp av 4-basskoden har DNA sekvensen för alla proteiner som finns i organismer, liksom element som reglerar deras uttryck..
DNA-strukturen måste vara stabil eftersom organismer använder den för att koda information. Det är dock en molekyl som är benägen för förändringar, så kallade mutationer. Dessa förändringar i det genetiska materialet är grundmaterialet för evolutionär förändring..
Liksom DNA är RNA en polymer av nukleotider, med undantaget att basen T ersätts med U. Denna molekyl är i form av ett enda band och uppfyller ett brett spektrum av biologiska funktioner..
I cellen finns tre huvud-RNA. Messenger RNA är en mellanhand mellan DNA och proteinbildning. Det ansvarar för att kopiera informationen i DNA: n och ta den till proteinöversättningsmaskineriet. Ribosomalt RNA, en andra typ, är en strukturell del av detta komplexa maskineri.
Den tredje typen, eller överför RNA, är ansvarig för att bära lämpliga aminosyrarester för syntes av proteiner.
Förutom de tre ”traditionella” RNA: erna finns det en serie små RNA som deltar i regleringen av genuttryck, eftersom i en cell kan alla gener som kodas i DNA inte uttryckas konstant och i samma storlek.
Organismer måste ha vägar för att reglera sina gener, det vill säga att avgöra om de uttrycks eller inte. På samma sätt består det genetiska materialet endast av en ordbok med spanska ord, och regleringsmekanismen möjliggör bildandet av ett litterärt verk.
Kvävebaser är en del av nukleosidtrifosfater, en molekyl som, liksom DNA och RNA, är av biologiskt intresse. Förutom basen består den av en pentos och tre fosfatgrupper kopplade samman genom högenergibindningar..
Tack vare dessa bindningar är nukleosidtrifosfater energirika molekyler och är huvudprodukten av metaboliska vägar som strävar efter att frigöra energi. Bland de mest använda är ATP.
ATP eller adenosintrifosfat består av den kvävehaltiga adeninen, kopplad till kolet placerat i position 1 i ett pentos-typ socker: ribos. I position fem av detta kolhydrat är de tre fosfatgrupperna kopplade.
I allmänhet är ATP cellens energivaluta, eftersom den kan användas och regenereras snabbt. Många vanliga metaboliska vägar bland organiska ämnen använder och producerar ATP.
Dess "kraft" är baserad på högenergibindningar, bildade av fosfatgrupper. De negativa laddningarna för dessa grupper är i konstant avstötning. Det finns andra orsaker som predisponerar hydrolys i ATP, inklusive resonansstabilisering och lösning.
Även om de flesta nukleosider saknar betydande biologisk aktivitet är adenosin ett tydligt undantag hos däggdjur. Detta fungerar som en autacoid, analog med ett "lokalt hormon" och som en neuromodulator.
Denna nukleosid cirkulerar fritt i blodomloppet och verkar lokalt med olika effekter på utvidgningen av blodkärlen, sammandragningar av glatt muskulatur, neuronal urladdning, frisättning av neurotransmittorn och fettmetabolism. Det är också relaterat till hjärtfrekvensreglering.
Denna molekyl är också involverad i regleringen av sömnmönster. Adenosinkoncentrationen ökar och främjar trötthet. Det är därför koffein hjälper oss att hålla oss vaken: det blockerar neurala interaktioner med extracellulärt adenosin..
Ett betydande antal vanliga metaboliska vägar i celler har regleringsmekanismer baserade på nivåerna av ATP, ADP och AMP. Dessa två sista molekyler har samma struktur som ATP, men har tappat en respektive två fosfatgrupper..
Som vi nämnde i föregående avsnitt är ATP en instabil molekyl. Cellen ska bara producera ATP när den behöver den, eftersom den måste använda den snabbt. ATP i sig är också ett element som reglerar metaboliska vägar, eftersom dess närvaro indikerar för cellen att det inte borde producera mer ATP..
Däremot varnar dess hydrolyserade derivat (AMP) cellen om att ATP tar slut och måste producera mer. Således aktiverar AMP metaboliska vägar för energiproduktion, såsom glykolys.
På liknande sätt medieras många hormonella signaler (såsom de som är involverade i glykogenmetabolism) intracellulärt av cAMP-molekyler (c är för cyklisk) eller av en liknande variant men med guanin i dess struktur: cGMP.
Vid flera steg i de metaboliska vägarna kan enzymer inte agera ensamma. De behöver ytterligare molekyler för att kunna utföra sina funktioner; Dessa element kallas koenzymer eller kosubstrat, varvid den senare termen är mer lämplig, eftersom koenzymer inte är katalytiskt aktiva.
I dessa katalytiska reaktioner finns det ett behov av att överföra elektronerna eller gruppen av atomer till ett annat substrat. Hjälpmolekylerna som deltar i detta fenomen är koenzymer.
Kvävebaser är strukturella element i dessa kofaktorer. Bland de mest kända är pyrimidinnukleotiderna (NAD+, NADP+), FMN, FAD och koenzym A. Dessa deltar i mycket viktiga metaboliska vägar, såsom glykolys, Krebs-cykeln, fotosyntes, bland andra.
Exempelvis är pyrimidinnukleotider mycket viktiga koenzymer av enzymer med dehydrogenasaktivitet och är ansvariga för transporten av hydridjoner..


Ingen har kommenterat den här artikeln än.