A kromosom duplicering beskriver en fraktion av DNA som visas två gånger som en produkt av genetisk rekombination. Kromosomal duplicering, genduplicering eller amplifiering är en av källorna till generationens variationer och evolution hos levande varelser.
En kromosomal duplicering är en typ av mutation, eftersom den innebär en förändring av den normala DNA-sekvensen i en kromosomregion. Andra mutationer på kromosomnivå inkluderar kromosomala insättningar, inversioner, translokationer och borttagningar..
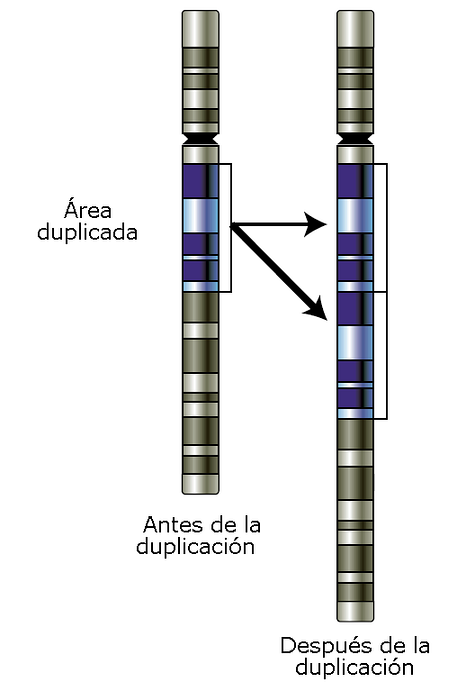
Kromosomala duplikationer kan förekomma på samma källplats som det duplicerade fragmentet. Dessa är batchdublikationerna. Partiduplikat kan vara av två typer: direkt eller inverterad.
Direktkopior är de som upprepar både informationen och riktningen för det upprepade fragmentet. I batch-inverterade duplikatfragment upprepas informationen, men fragmenten är orienterade i motsatta riktningar.
I andra fall kan kromosomal duplicering inträffa på en annan plats eller till och med på en annan kromosom. Detta genererar en ektopisk kopia av sekvensen som kan fungera som ett substrat för crossover och vara en källa till avvikande rekombinationer. Beroende på storleken som är inblandad kan dupliceringarna vara makro- eller mikroduplikationer.
Evolutionärt sett genererar duplikationer variation och förändring. På individnivå kan emellertid kromosomala duplikationer leda till allvarliga hälsoproblem..
Artikelindex
Duplikationer förekommer oftast i DNA-områden som har upprepade sekvenser. Dessa är substrat för rekombinationshändelser, även om de förekommer mellan regioner som inte är helt homologa..
Dessa rekombinationer sägs vara olagliga. Mekaniskt beror de på sekvenslikhet, men genetiskt kan de utföras mellan icke-homologa kromosomer.
Hos människan har vi flera typer av repetitiva sekvenser. De mycket repetitiva inkluderar så kallat satellit-DNA, begränsat till centromererna (och vissa heterokromatiska regioner).
Andra, måttligt repetitiva, inkluderar till exempel satsen som upprepar den kod för ribosomala RNA. Dessa upprepade eller duplicerade regioner finns i mycket specifika platser som kallas nukleoliorganisationsregioner (NOR)..
NOR, hos människor, ligger i de subtelomera regionerna i fem olika kromosomer. Varje NOR består å sin sida av hundratals till tusentals kopior av samma kodningsregion i olika organismer..
Men vi har också andra repetitiva regioner utspridda genom genomet, med olika sammansättning och storlekar. Alla kan rekombineras och ge upphov till duplikationer. Faktum är att många av dem är produkten av sin egen duplicering, in situ eller ektopisk. Dessa inkluderar, men är inte begränsade till, minisatelliter och mikrosatelliter.
Kromosomala duplikationer kan också uppstå, mer sällan, genom sammanfogning av icke-homologa ändar. Detta är en icke-homolog rekombinationsmekanism som observeras i vissa reparationshändelser med dubbla band i DNA..
När en gen dupliceras på samma plats, eller till och med på en annan, skapar den en plats med sekvens och betydelse. Det vill säga en meningsfull sekvens. Om det förblir så kommer det att vara en duplikatgen av och från dess modergen..
Men det kan hända att det inte utsätts för samma selektiva tryck som modergenen och kan mutera. Summan av dessa ändringar kan ibland leda till att en ny funktion visas. Genen kommer också att vara en ny gen.
Kopiering av det förfädernas globinlokal, till exempel, ledde i evolutionen till framväxten av globinfamiljen. Efterföljande translokationer och efterföljande duplikationer fick familjen att växa med nya medlemmar som utför samma funktion, men lämpar sig för olika förhållanden.
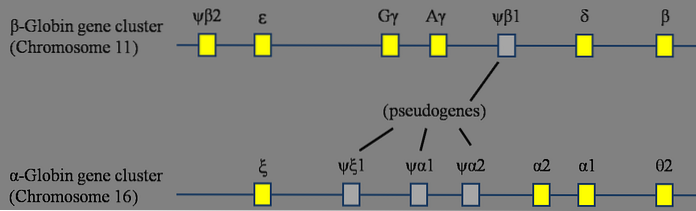
I en organism leder duplicering av en gen till genereringen av en kopia som kallas en paraloggen. Ett välstuderat fall är det av de globingener som nämns ovan. En av de mest kända globinerna är hemoglobin.
Det är mycket svårt att föreställa sig att endast den kodande regionen för en gen dupliceras. Därför är varje paraloggen associerad med en paralogregion i organismen som genomgår dubbelarbete..
Under utvecklingen har kromosomala duplikationer spelat en viktig roll på olika sätt. Å ena sidan duplicerar de informationen som kan ge upphov till nya funktioner på grund av förändringar i gener med tidigare funktion..
Å andra sidan kan placera duplicering i ett annat genomiskt sammanhang (till exempel en annan kromosom) generera en paralog med olika reglering. Det vill säga det kan generera större anpassningsförmåga.
Slutligen skapas utbytesregioner också genom rekombination som leder till stora genomiska omläggningar. Detta kan i sin tur representera ursprunget till specieringshändelser i särskilt makroevolutionära linjer..
Framsteg inom nästa generations sekvenseringsteknik, såväl som kromosomfärgning och hybridisering, gör att vi nu kan se nya föreningar. Dessa föreningar inkluderar manifestationen av vissa sjukdomar på grund av vinst (duplicering) eller förlust (radering) av genetisk information..
Genetiska duplikationer är associerade med en förändring i gendosering och med avvikande övergångar. I vilket fall som helst leder de till en obalans mellan genetisk information, som ibland manifesterar sig som en sjukdom eller ett syndrom..
Charcot-Marie-Tooth syndrom typ 1A är till exempel associerad med mikroduplicering av regionen som inkluderar PMP22-genen. Syndromet är också känt under namnet ärftlig motorisk och sensorisk neuropati.
Det finns kromosomfragment som är benägna att dessa förändringar. Faktum är att 22q11-regionen har många låga kopieringsantal som är specifika för den delen av genomet..
Det vill säga från regionen av band 11 i den långa armen av kromosom 22. Dessa duplikationer är associerade med många genetiska störningar, inklusive mental retardation, missbildningar i ögonen, mikrocefali, etc..
I fall av mer omfattande duplikationer kan partiella trisomier förekomma med skadliga effekter på organismens hälsa..


Ingen har kommenterat den här artikeln än.