De isogami det är ett växtreproduktionssystem där könscellerna är morfologiskt lika. Likhet förekommer i form och storlek, och manliga och kvinnliga könsceller kan inte urskiljas. Detta reproduktionssystem anses vara förfäder. Det förekommer i olika grupper av alger, svampar och protozoer.
De könsceller som är inblandade i isogami kan vara mobila (ciliated) eller inte. Deras förening sker genom konjugation. Odifferentierade könsceller smälter och utbyter genetiskt material.
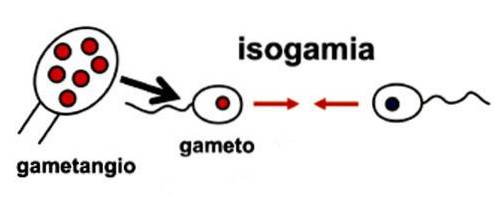
Isogami kan vara homotisk eller heterotisk. Det är homotiskt när fusionen sker mellan könsceller som har samma genom. I heterotal isogami har könsceller en annan genetisk sammansättning.
Artikelindex
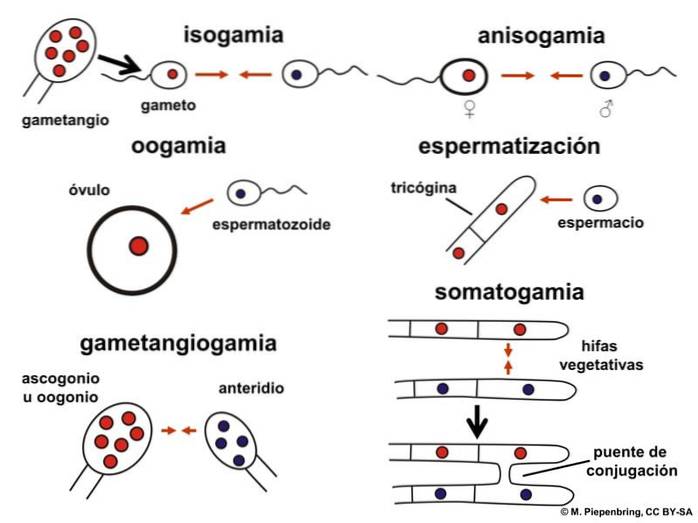
Reproduktion med isogami sker genom konjugering. I detta rör sig innehållet i en cell mot en annan och fusion uppstår.
Processer av karigami (fusion av kärnor) och plasmogami (fusion av cytoplasma) är inblandade. Somatisk celldifferentiering till sexuella celler kan associeras med miljöförhållanden. Interaktion med andra individer av samma art kan också påverka.
Efter att differentiering inträffat måste könscellerna hitta och känna igen andra könsceller. I grupper där isogami förekommer sker igenkänning och fusion av könsceller på olika sätt.
Sexceller kan flagelleras eller orörliga. I vissa fall är de stora, som i vissa gröna alger.
Det finns två typer av isogami relaterade till den genetiska sammansättningen av könsceller.
En individs gamete är konjugerad med en annan från samma klongrupp. I detta fall anses självbefruktning förekomma.
Alla kärnor har samma genotyp och det finns ingen interaktion med en annan genotyp. Somatiska celler differentieras direkt till könsceller.
Könsceller bildas i klonala populationer och senare fusion uppstår för att bilda zygoten.
Gameter produceras i olika individer, som har olika genetisk sammansättning.
Könsceller måste ha genetisk kompatibilitet för att fusion ska uppstå. Två typer av gameter bildas generellt. "Plus" och "minus" som är kompatibla med varandra.
Gametangialcellen (som producerar gameten) av en typ bildar ett par med den för den andra typen. Dessa känns igen genom kemisk kommunikation som i vissa fall involverar feromonproduktion.
Isogamins tillstånd verkar dominera i encelliga organismer, medan anisogami är nästan universellt för multicellulära eukaryoter. I de flesta eukaryota släkter av encelliga organismer är könscellerna lika stora och vi skiljer inte mellan män och kvinnor..
I eukaryoter finns det ett betydande antal arter med isogamiska könsceller. Vi kommer dock bara att nämna de släktingar som uppträder ständigt i den biologiska litteraturen - även om det finns många fler..
Den välkända sociala amöben av arten Dictyostelium discoideum, den vanliga jästen vi använder för att göra mat Saccharomyces cerevisiae och den protozoiska parasiten som orsakar sömnsjuka Trypanosoma brucei är alla exempel på organismer med identiska könsceller.
I gröna alger är isogami ett vanligt fenomen. I själva verket finns det två typer av isogami i dessa organismer.
Vissa arter producerar relativt medelstora könsceller med ett fototaktiskt system som representeras av en ögonfläck. Andra arter har samma könsceller, men mycket mindre än i föregående fall. De saknar också ögonfläck.
Det är dock inte möjligt att göra en sådan radikal observation och begränsa isogamiska gameter till encelliga linjer och anisogamic till multicellular varelser..
Faktum är att växter utgör några undantag från denna regel, eftersom koloniala gröna alger som t.ex. Pandorina, Volvulina Y Yamagishiella presentera villkoret för isogami.
Det finns också undantag i motsatt riktning, eftersom det finns encelliga organismer, såsom de gröna algerna av ordningen Bryopsidales som presenterar olika könsceller..
I alger har närvaron av två typer av könsceller associerade med isogami observerats.
I vissa grupper är könscellerna medelstora och har fototaxmekanismer. Det finns en ögonfläck som stimuleras av ljus.
De är vanligtvis förknippade med närvaron av kloroplaster och förmågan att ackumulera reservämnen. I andra fall är könscellerna mycket små och har ingen ögonfläck..
Sexuell reproduktion hos isogami-alger sker annorlunda.
Det är en grupp unicellulära gröna alger med två flageller. Den presenterar heterotisk isogami. Homotisk isogami kan förekomma hos vissa arter.
Haploida vegetativa celler differentieras till könsceller när kväveförhållandena ökar i mediet. Det finns två typer av könsceller, med olika genetiska komplement.
Gameter producerar agglutininer (vidhäftningsmolekyler) som främjar bindningen av flageller. Efter fusion tillhandahåller de två könscellerna den genetiska information som är nödvändig för embryonets utveckling.
Dessa alger tillhör Charyophyta-divisionen. De är unicellular. De presenterar homotisk och heterotisk isogami.
Gameter är inte mobila. I det här fallet, när könscellerna har sitt ursprung, bildas en konjugeringspapill. Cytoplasmer frigörs genom bristning på cellväggen.
Därefter sker sammansmältningen av protoplasmerna i båda könscellerna och zygoten bildas. Kemisk attraktion mellan de olika genetiska typerna anses förekomma i heterotisk isogami.
De är flercelliga organismer, med isogamiska flagellater. Andra grupper reproducerar med anisogami eller oogamy.
Gameter är morfologiskt desamma, men de beter sig annorlunda. Det finns arter där den kvinnliga typen släpper feromoner som lockar den manliga typen.
I andra fall rör sig en typ av gamet under en kort period. Ta sedan in flagellen och släpp feromoner. Den andra typen rör sig under längre tid och har en feromonsignalreceptor.
Både homotala och heterotiska typer är isogami. I de flesta fall är könsigenkänning associerad med produktion av feromoner.
I olika encelliga grupper såsom Saccharomyces, könsceller differentierar som svar på en förändring av odlingsmediets sammansättning. Somatiska celler delar sig med meios under vissa förhållanden, såsom låga kvävehalter..
Gameter med olika genetiska sammansättningar känns igen av feromonsignaler. Cellerna bildar utsprång mot feromonkällan och sammanfogar deras toppar. Kärnorna i båda könscellerna migrerar tills de smälter samman och bildar en diploid cell (zygote).
De är flercelliga organismer. De presenterar främst heterotiska system. Under sexuell utveckling bildar de donator- (manliga) och mottagliga (kvinnliga) strukturer.
Cellfusion kan förekomma mellan en hyfa och en mer specialiserad cell eller mellan två hyfer. Inmatningen av givarkärnan (han) i hyfa stimulerar utvecklingen av en fruktkropp.
Kärnor smälter inte omedelbart. Fruktkroppen bildar en dikaryot struktur med kärnor med olika genetisk sammansättning. Därefter smälter och delar kärnorna med meios.
Isogamy förekommer i flagellat encelliga grupper. Dessa cilierade organismer etablerar cytoplasmatisk koppling mellan könsceller i specialområden av plasmamembranet..
De cilierade grupperna har två kärnor, en makronukleus och en mikronukleus. Makronkärnan är den somatiska formen. Den diploida mikrokärnan delar med meios och bildar könscellen.
De haploida kärnorna utbyts av en cytoplasmatisk brygga. Därefter återställs cytoplasman i varje cell och de återfår sin autonomi. Denna process är unik inom eukaryoter.
På Euplots Specifika feromoner av varje genetisk typ produceras. Celler stoppar somatisk tillväxt när de upptäcker en feromon med olika genetisk sammansättning.
För arter av Dileptus igenkänningsmolekyler presenteras på cellytan. Kompatibla gameter är bundna av vidhäftningsproteiner i cilier.
På Paramecium igenkänningsämnen produceras mellan kompatibla könsceller. Dessa ämnen främjar förening av könsceller, liksom deras vidhäftning och efterföljande fusion..
I evolutionärbiologi är ett av de mest diskuterade ämnena när vi pratar om komplexa organismer (som däggdjur) föräldrarnas investeringar. Detta koncept utvecklades av den framstående biologen Sir Ronald Fisher i sin bok "Den genetiska teorin om naturligt urval”, Och innebär föräldrarnas kostnader för avkommans välbefinnande.
Jämställdhet i könsorgan innebär att föräldrarnas investeringar kommer att vara symmetriska för båda organismerna som är involverade i den reproduktiva händelsen..
Till skillnad från anisogamisystemet, där föräldrarnas investeringar är asymmetriska, och det är den kvinnliga könscellen som ger de flesta icke-genetiska resurser (näringsämnen, etc.) för zygotutveckling. Med utvecklingen av system som presenterar dimorfism i sina gameter utvecklades också en asymmetri i föräldrarnas organismer.
Enligt bevisen och reproduktionsmönstren som vi hittar hos moderna arter verkar det logiskt att betrakta isogami som det förfädernas tillstånd, som uppträder i de första stadierna av sexuell reproduktion..
I flera släkter av flercelliga organismer, såsom växter och djur, har ett differentiellt reproduktionssystem utvecklats oberoende, där de kvinnliga könscellerna är stora och orörliga och de manliga är små och med förmågan att flytta till ägglossningen..
Även om de exakta banorna för förändring från ett isogamiskt till ett anisogamiskt tillstånd inte är kända, har flera teorier formulerats..
En av dem belyser en möjlig avvägning mellan storleken på könscellerna och deras antal. Enligt detta argument är anisogamins ursprung en evolutionärt stabil strategi orsakad av störande urval i sökandet efter zygotens effektivitet och överlevnad..
En annan teori försöker förklara fenomenet som ett sätt att kompensera för en orörlig cell (äggstocken) med många celler med förmåga att röra sig (spermierna)..
En tredje vy förklarar genereringen av anisogami som en adaptiv egenskap för att undvika konflikter mellan kärnan och cytoplasman på grund av organellernas uniparental arv..


Ingen har kommenterat den här artikeln än.