De prometafas det är ett stadium av celldelningsprocessen, mellanliggande mellan profas och metafas. Det kännetecknas av interaktionen mellan de delande kromosomerna och mikrotubuli som är ansvariga för att separera dem. Prometafas förekommer i både mitos och meios, men med olika egenskaper.
Det tydliga målet för all celldelning är att producera fler celler. För att uppnå detta måste cellen ursprungligen duplicera sitt DNA-innehåll; replikera det. Utöver detta måste cellen separera dessa kromosomer på ett sådant sätt att det specifika syftet med varje uppdelning av cytoplasman uppfylls..

Vid mitos, samma antal kromosomer från modercellen i dottercellerna. I meios I, separationen mellan homologa kromosomer. I meios II, separationen mellan systerkromatider. Det är, i slutet av processen, få de fyra förväntade meiotiska produkterna.
Cellen hanterar denna komplicerade mekanism genom användning av specialkomponenter som mikrotubuli. Dessa är organiserade av centrosomen i de flesta eukaryoter. I andra, tvärtom, såsom högre växter, fungerar en annan typ av organisationscentrum för mikrotubuli.
Artikelindex

Mikrotubuli är linjära polymerer av proteinet tubulin. De är involverade i nästan alla cellulära processer som involverar förskjutning av någon intern struktur. De är en integrerad del av cytoskelettet, cilia och flagella.
När det gäller växtceller spelar de också en roll i den interna strukturella organisationen. I dessa celler bildar mikrotubuli ett slags gobeläng fäst vid plasmamembranets inre yta..
Denna struktur, som kontrollerar växtcelldelningar, är känd som den kortikala organisationen av mikrotubuli. Vid tidpunkten för mitotisk uppdelning kollapsar de till exempel i en central ring som kommer att vara den framtida platsen för den centrala plattan, i planet där cellen kommer att dela sig..
Mikrotubuli består av alfa-tubulin och beta-tubulin. Dessa två underenheter bildar en heterodimer, som är den grundläggande strukturella enheten för tubulinfilament. Polymerisationen av dimererna leder till bildandet av 13 protofilament i en lateral organisation som ger upphov till en ihålig cylinder.

De ihåliga cylindrarna med denna struktur är mikrotubuli, som genom sin egen sammansättning visar polaritet. Det vill säga den ena änden kan växa genom tillsats av heterodimerer, medan den andra kan genomgå subtraktioner. I det senare fallet krymper röret i stället för att förlängas i den riktningen.
Mikrotubuli kärnbildar (det vill säga börjar polymerisera) och organiseras i mikrotubuli-organiseringscentra (COM). COM är associerat med centrosomer under uppdelningar i djurceller.
I högre växter, som inte har centrosomer, finns COM på analoga platser, men består av andra komponenter. I cilia och flagella är COM basal mot motorstrukturen.
Förskjutningen av kromosomer under celldelning uppnås genom mikrotubuli. Dessa förmedlar den fysiska interaktionen mellan kromosomernas centromerer och COM.
Genom riktade depolymeriseringsreaktioner kommer metafaskromosomer så småningom att röra sig mot polerna i delande celler..
Korrekt mitotisk kromosomavskiljning är en som garanterar att varje dottercell får ett komplement av kromosomer som är identiska med modercellens..
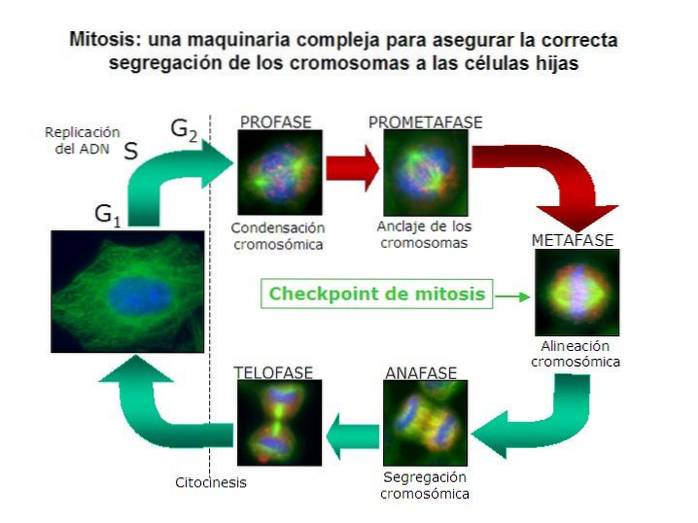
Detta innebär att cellen måste separera varje par duplicerade kromosomer i två separata, individuella kromosomer. Det vill säga det måste separera systerkromatiderna för varje homologt par från hela komplementet av stamcellens kromosomer..
I öppna mitoser är försvinnandet av kärnhöljet kännetecknet för prometafas. Detta gör att det enda hindret mellan MOC och kromosomernas centromer försvinner..
Från MOC: erna polymeriseras långa mikrotubuli-filament och förlängs mot kromosomerna. När man hittar en centromer upphör polymerisationen och en kromosom fäst vid en COM erhålls..
I mitos är kromosomerna dubbla. Därför finns det också två centromerer, men fortfarande förenade i samma struktur. Detta betyder att i slutet av mikrotubuli-polymerisationsprocessen kommer vi att ha två av dem per duplicerad kromosom..
En glödtråd fäster en centromer till en COM, och en annan till systerkromatiden som är fäst vid COM motsatt den första..
I slutna mitoser är processen nästan identisk med den tidigare, men med stor skillnad; kärnkraftshöljet försvinner inte. Därför är COM internt och är associerat med det interna kärnkraftshöljet genom kärnplattan..
I halvstängda (eller halvöppna) mitoser försvinner kärnhöljet bara vid de två motsatta punkterna där en mitotisk COM existerar utanför kärnan..
Detta innebär att mikrotubuli i dessa mitoser tränger in i kärnan för att kunna mobilisera kromosomerna i steg efter prometafaset..
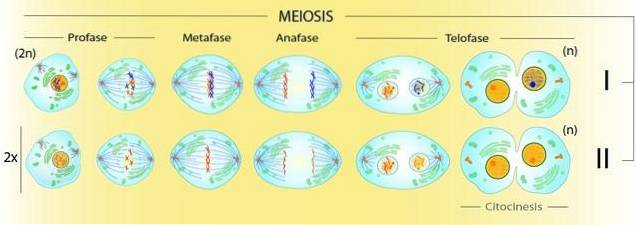
Eftersom meios involverar produktion av fyra 'n' celler från en '2n' cell måste det finnas två delar av cytoplasman. Låt oss titta på det så här: i slutet av metafas I kommer det att finnas fyra gånger fler kromatider än centromerer synliga under mikroskopet.
Efter den första uppdelningen kommer det att finnas två celler med dubbelt så många kromatider som centromerer. Först i slutet av den andra cytoplasmiska uppdelningen kommer alla centromerer och kromatider att individualiseras. Det kommer att finnas lika många centromerer som det finns kromosomer.
Nyckelproteinet för att dessa komplexa interkromatininteraktioner ska äga rum vid mitos och meios är kohesin. Men det finns fler komplikationer i meios än vid mitos. Det är därför inte förvånande att meiotisk kohesin skiljer sig från mitotisk.
Kohesiner möjliggör sammanhållning av kromosomer under deras mitotiska och meiotiska kondensprocess. Dessutom tillåter de och reglerar interaktionen mellan systerkromatider i båda processerna..
Men i meios främjar de också något som inte förekommer vid mitos: homolog parning och de därav följande synapserna. Dessa proteiner är olika i båda fallen. Vi kan säga att meios utan en sammanhållning som skiljer den inte skulle vara möjlig.
Mekaniskt sett är centromeren / COM-interaktionen densamma i all celldelning. Men i prometafas I av meios I skiljer cellen inte systerkromatider som det gör vid mitos..
Däremot har den meiotiska tetrad fyra kromatider i en uppenbar dubbel uppsättning centromerer. I denna struktur finns det något annat som inte finns i mitos: chiasmata.
Kiasmer, som är fysiska korsningar mellan homologa kromosomer, skiljer vilka centromerer som måste separeras: de av homologa kromosomer.
Således bildas i prometafas I kopplingar mellan homologernas centromerer och COM vid motsatta poler i cellen..
Detta prometafas II liknar mer mitotiskt prometafas än meiotiskt prometafas I. I det här fallet kommer COM: er att "lansera" mikrotubuli vid de duplicerade centromererna av systerkromatider..
Således kommer två celler att produceras med individuella kromosomer, produkten av en kromatid från varje par. Därför kommer celler med det haploida kromosomkomplementet som arten ges.

Ingen har kommenterat den här artikeln än.