
De dihybridism, I genetik definierar den samtidig studie av två olika ärftliga karaktärer och i förlängning av dem vars manifestation beror på två olika gener, även om det har samma karaktär.
De sju drag som Mendel analyserade var användbara för honom när han lade fram sin teori om karaktärsarv eftersom bland annat generna som var ansvariga för deras manifestation hade kontrasterande alleler vars fenotyp var lätt att analysera och eftersom var och en bestämde uttrycket enda karaktär.
Det vill säga de var monogena egenskaper vars hybridtillstånd (monohybrider) gjorde det möjligt att bestämma dominans / recessivitetsförhållandena mellan allelerna för den enskilda genen..
När Mendel analyserade det gemensamma arvet av två olika karaktärer, fortsatte han som han gjorde med enstaka karaktärer. Han fick dubbla hybrider (dihybrider) som gjorde det möjligt för honom att kontrollera:
Vi vet nu att arv av karaktärer är lite mer komplicerat än vad Mendel observerade, men också att Mendel i grunden var helt korrekt..
Den efterföljande utvecklingen av genetik gjorde det möjligt att visa att dihybridkorsningar och deras analys (dihybridism), som Bateson ursprungligen kunde demonstrera, kunde vara en outtömlig källa till upptäckter i denna kraftfulla och framväxande vetenskap på 1900-talet..
Tack vare deras smarta användning kunde de ge genetikern en något tydligare uppfattning om beteendet och naturen hos gener..
Artikelindex
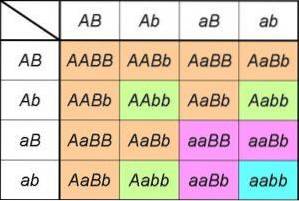
Om vi analyserar produkterna från ett monohybridkors Aa X Aa, vi kan märka att det är lika med att utveckla den anmärkningsvärda produkten (TILL+till)två= AA + 2Aa + aa.
Uttrycket till vänster inkluderar de två typerna av könsceller som en av föräldrarna som är heterozygota för genen kan producera TILL/till; genom att kvadrera indikerar vi att båda föräldrarna har samma sammansättning för genen som studeras [det vill säga det är ett monohybridkors (TILL+till) X (TILL+till)].
Uttrycket till höger ger oss genotyperna (och därför härleds fenotyperna) och förväntade proportioner härledda från korsningen.
Därför kan vi direkt observera de genotypiska proportionerna härledda från den första lagen (1: 2: 1), liksom de fenotypiska proportionerna som förklaras av den (1AA+tvåAa= 3TILL_ för varje 1aa, eller fenotypiskt förhållande 3: 1).
Om vi nu överväger ett kors för att analysera arv av en gen B, Uttrycken och proportionerna kommer att vara desamma; i själva verket kommer det att vara så för någon gen. I ett dihybridkors har vi därför utvecklingen av produkterna från (TILL+till)två X (B+b)två.
Eller vad är detsamma, om dihybridkorset involverar två gener som deltar i arv av två orelaterade karaktärer, kommer de fenotypiska proportionerna att vara de som förutses av den andra lagen: (3TILL_: 1aa) X (3B_: 1bebis) = 9TILL_B_: 3TILL_bebis: 3aaB_: 1aabb).
Dessa härleds naturligtvis från de ordnade genotypförhållandena 4: 2: 2: 2: 2: 1: 1: 1: 1 som härrör från produkten från (TILL+till)två X (B+b)två= (AA + 2Aa + aa) X (BB + tvåBb + bebis).
Vi inbjuder dig att se för dig själv för att nu analysera vad som händer när de fenotypiska förhållandena 9: 3: 3: 1 i ett dihybridkors "avviker" från dessa tydliga och förutsägbara matematiska förhållanden som förklarar det oberoende arvet av två kodade tecken. olika gener.
Det finns två huvudsakliga sätt som dihybridkorsningar avviker från vad som förväntas. Den första är den där vi analyserar det gemensamma arvet av två olika karaktärer, men de fenotypiska proportionerna som observeras i avkomman ger en tydlig övervägande för manifestationen av föräldrafenotyper..
Det är troligtvis ett fall av länkade gener. Det vill säga, de två gener som analyseras, även om de befinner sig på olika platser, är så nära varandra fysiskt att de tenderar att ärvas tillsammans och uppenbarligen distribueras de inte oberoende av varandra..
Den andra omständigheten, som också är ganska vanlig, härrör från det faktum att en liten minoritet av ärftliga egenskaper är monogena..
Tvärtom deltar mer än två gener i manifestationen av de flesta ärftliga egenskaper..
Av denna anledning är det alltid möjligt att de genetiska interaktioner som upprättas mellan generna som deltar i manifestationen av en enda karaktär är komplexa och går utöver ett enkelt förhållande av dominans eller recessivitet som observerats i relationer alleliska typiska för monogena egenskaper.
Till exempel i manifestationen av ett drag kan cirka fyra enzymer delta i en viss ordning för att ge upphov till den slutliga produkten som är ansvarig för den fenotypiska manifestationen av den vilda fenotypen..
Analysen som gör det möjligt att identifiera antalet gener från olika loci som deltar i manifestationen av en genetisk egenskap, liksom i vilken ordning de verkar, kallas epistasanalys och är kanske den som oftast definierar vad vi kallar genetisk analys. i sin mest klassiska mening.
I slutet av detta inlägg presenteras de fenotypiska proportionerna som observeras i de vanligaste fallen av epistasis - och detta tar endast hänsyn till dihybridkorsningar..
Genom att öka antalet gener som deltar i manifestationen av samma drag ökar naturligtvis komplexiteten av geninteraktioner och deras tolkning..
Dessutom, som i sin tur kan tas som den gyllene regeln för en korrekt diagnos av epistatiska interaktioner, kan utseendet på nya fenotyper som inte finns i föräldragenerationen verifieras..
Slutligen, förutom att tillåta analys av utseendet på nya fenotyper och deras andel, tillåter analysen av epistasis också att bestämma den hierarkiska ordning i vilken de olika generna och deras produkter måste manifestera sig i en given väg för att redogöra för den fenotyp som är associerad med dem..
Den mest grundläggande eller tidiga manifestationsgenen är epistatisk över alla andra, eftersom utan dess produkt eller verkan, till exempel, kommer de nedströms om den inte att kunna uttrycka sig, vilket därför kommer att vara hypostatiskt för den..
En gen / produkt på tredje plats i hierarkin kommer att vara hypostatisk mot de två första och epistatisk för alla andra som finns kvar i denna genuttrycksväg..


Ingen har kommenterat den här artikeln än.