
De endoplasmatiska retiklet det är en membranös cellulär organell som finns i alla eukaryota celler. Detta komplexa system upptar ungefär mer än hälften av membranen i en vanlig djurcell. Membranen fortsätter tills de möter kärnmembranet och bildar ett kontinuerligt element.
Denna struktur fördelas genom cellcytoplasman i form av en labyrint. Det är ett slags nätverk av tubuli kopplade till varandra med säckliknande strukturer. Inom endoplasmatisk retikulum förekommer protein- och lipidbiosyntes. Nästan alla proteiner som måste transporteras till utsidan av cellen passerar först genom nätverket.
Retikulummembranet är inte bara ansvarigt för att separera det inre av denna organell från det cytoplasmiska utrymmet och förmedla transporten av molekyler mellan dessa cellfack; Det är också involverat i syntesen av lipider, som kommer att utgöra en del av plasmamembranet i cellen och membranet i de andra organellerna..
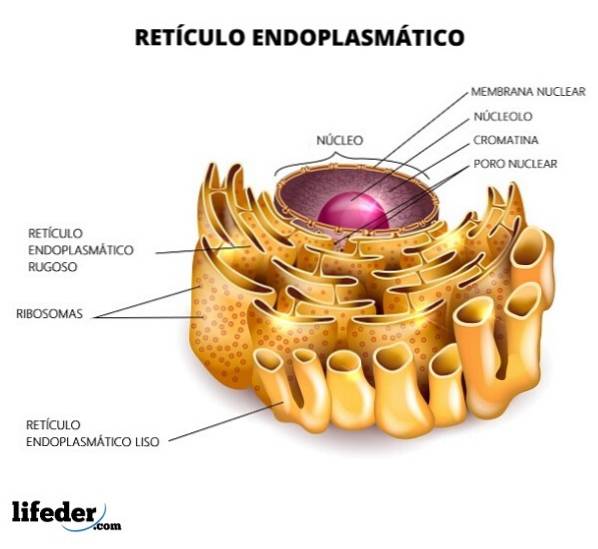
Retikulumet är uppdelat i slät och grov, beroende på närvaron eller frånvaron av ribosomer i dess membran. Det grova endoplasmiska retikulumet har ribosomer fästa vid membranet (närvaron av ribosomer ger det ett "grovt" utseende) och tubulärformen är något rak.
För sin del saknar det släta endoplasmiska retikulumet ribosomer och strukturen är mycket mer oregelbunden. Funktionen hos det grova endoplasmatiska retikulumet är främst inriktat på bearbetning av proteiner. Däremot är smooth ansvarig för lipidmetabolism.
Artikelindex
Det endoplasmiska retikulumet är ett membranöst nätverk som finns i alla eukaryota celler. Den består av saccules eller cisterner och rörformiga strukturer som bildar ett kontinuum med kärnans membran och är fördelade över cellen.
Retikulumlumen kännetecknas av höga koncentrationer av kalciumjoner, förutom en oxiderande miljö. Båda egenskaperna gör att den kan fullgöra sina funktioner.
Det endoplasmiska retikulumet anses vara den största organellen som finns i celler. Cellvolymen i detta fack täcker cirka 10% av cellens inre.
Det grova endoplasmiska retikulumet har en hög densitet av ribosomer på ytan. Det är regionen där alla processer relaterade till proteinsyntes och modifiering sker. Dess utseende är främst rörformigt.
Det släta endoplasmiska retikulumet har inte ribosomer. Det finns rikligt i celltyper som har en aktiv metabolism i lipidsyntes; till exempel i cellerna i testiklarna och äggstockarna, som är steroidproducerande celler.
På liknande sätt finns det släta endoplasmatiska retikulumet i en ganska hög andel i leverceller (hepatocyter). Produktionen av lipoproteiner sker i detta område..
Jämfört med det grova endoplasmiska nätverket är dess struktur mer komplicerad. Överflödet av det släta kontra grova nätverket beror främst på celltypen och dess funktion..
Den fysiska arkitekturen för det endoplasmiska nätverket är ett kontinuerligt system av membran som består av sammankopplade säckar och tubuli. Dessa membran sträcker sig till kärnan och bildar ett enda lumen.
Gitteret är byggt av flera domäner. Fördelningen är associerad med andra organeller, olika proteiner och komponenter i cytoskelettet. Dessa interaktioner är dynamiska.
Strukturellt består det endoplasmiska retikulumet av kärnhöljet och det perifera endoplasmiska retikulumet, som består av tubuli och säckar. Varje struktur är relaterad till en specifik funktion.
Kärnhöljet, som alla biologiska membran, består av ett lipid dubbelskikt. Interiören avgränsad av detta delas med det perifera nätverket.
Säckarna som utgör det endoplasmiska nätverket är plana och ofta staplade. De innehåller böjda områden vid membranens kanter. Det rörformiga nätverket är inte en statisk enhet; kan växa och omstrukturera.
Säck- och tubuli-systemet finns i alla eukaryota celler. Det varierar dock i form och struktur beroende på celltyp..
Retikulumet för celler med viktiga funktioner i proteinsyntes består huvudsakligen av säckar, medan cellerna som är mest relaterade till lipidsyntes och kalciumsignalering består av ett större antal tubuli..
Exempel på celler med ett stort antal säckar är de sekretoriska cellerna i bukspottkörteln och B-cellerna. Däremot har muskelceller och leverceller ett nätverk av framstående tubuli..
Det endoplasmiska retikulumet är involverat i ett antal processer inklusive proteinsyntes, handel och vikning och modifieringar, såsom bildning av disulfidbindning, glykosylering och tillsats av glykolipider. Dessutom deltar den i biosyntesen av membranlipider.
Nya studier har relaterat nätverket till svar på cellulär stress och kan till och med inducera apoptosprocesser, även om mekanismerna inte har klarlagts helt. Alla dessa processer beskrivs i detalj nedan:
Det endoplasmiska retikulumet är nära kopplat till proteinhandel; specifikt till proteiner som måste skickas utomlands, till Golgi-apparaten, till lysosomer, till plasmamembranet och logiskt sett till de som tillhör samma endoplasmatiska retikulum.
Det endoplasmiska retikulum är det cellulära beteende som är involverat i syntesen av proteiner som måste bäras utanför cellen. Denna funktion belystes av en grupp forskare på 1960-talet och studerade bukspottkörtelns celler vars funktion är att utsöndra matsmältningsenzymer..
Denna grupp, ledd av George Palade, lyckades märka proteiner med radioaktiva aminosyror. På detta sätt var det möjligt att spåra och lokalisera proteinerna med en teknik som kallas autoradiografi..
Radioaktivt märkta proteiner kan spåras till det endoplasmiska retikulumet. Detta resultat indikerar att retikulum är involverat i syntesen av proteiner vars slutdestination är utsöndring..
Senare flyttas proteinerna till Golgi-apparaten, där de "packas" i blåsor vars innehåll kommer att utsöndras..
Sekretionsprocessen inträffar eftersom blåsornas membran kan smälta samman med plasmamembranet i cellen (båda är lipid till sin natur). På detta sätt kan innehållet släppas på utsidan av cellen..
Med andra ord måste utsöndrade proteiner (och även proteiner riktade mot lysosomer och plasmamembranet) följa en specifik väg som involverar det grova endoplasmatiska retikulumet, Golgi-apparaten, sekretoriska vesiklar och slutligen cellens utsida..
Proteiner som är avsedda att införlivas i något biomembran (plasmamembran, Golgi-membran, lysosom eller retikulum) införs först i retikulamembranet och släpps inte direkt i lumen. De måste följa samma väg för sekretoriska proteiner.
Dessa proteiner kan lokaliseras i membranen genom en hydrofob sektor. Denna region har en serie av 20 till 25 hydrobofiska aminosyror, som kan interagera med koldioxidkedjorna i fosfolipider. Emellertid är det sätt på vilket dessa proteiner sätts in varierande..
Många proteiner passerar membranet bara en gång, medan andra gör det flera gånger. På samma sätt kan det i vissa fall vara slutet på karboxylterminalen eller aminoterminalen.
Orienteringen av nämnda protein fastställs medan peptiden växer och överförs till det endoplasmiska retikulumet. Alla proteindomäner som pekar mot retikulumlumen finns på utsidan av cellen i sin slutliga position..
Molekyler av proteinkaraktär har en tredimensionell konformation som är nödvändig för att utföra alla sina funktioner..
DNA (deoxiribonukleinsyra), genom en process som kallas transkription, överför sin information till en RNA (ribonukleinsyra) -molekyl. RNA passerar sedan in i proteinerna genom translationen. Peptider överförs till nätverket när översättningen pågår.
Dessa kedjor av aminosyror är ordnade på ett tredimensionellt sätt i retikulum med hjälp av proteiner som kallas chaperoner: ett protein från Hsp70-familjen (värmechockproteiner eller värmechockproteiner för dess förkortning på engelska; siffran 70 hänvisar till dess atommassa, 70 KDa) kallad BiP.
BiP-proteinet kan binda till polypeptidkedjan och förmedla dess vikning. På samma sätt deltar den i sammansättningen av de olika underenheterna som utgör proteinernas kvaternära struktur..
Proteiner som inte har vikts korrekt behålls av retikulum och förblir bundna till BiP eller bryts ned.
När cellen utsätts för påfrestningsförhållanden reagerar näthinnan på den och följaktligen sker inte korrekt veckning av proteiner. Cellen kan vända sig till andra system och producera proteiner som upprätthåller retikulumhomeostas.
En disulfidbrygga är en kovalent bindning mellan sulfhydrylgrupperna som ingår i aminosyrastrukturen cystein. Denna interaktion är avgörande för att vissa proteiner fungerar; på samma sätt definierar den strukturen hos proteinerna som presenterar dem.
Dessa bindningar kan inte bildas i andra cellfack (till exempel i cytosolen), eftersom den inte har en oxiderande miljö som gynnar dess bildning..
Det finns ett enzym involverat i bildandet (och brytningen) av dessa bindningar: proteindisulfidisomeraset.
I retikulum sker glykosyleringsprocessen vid specifika asparaginrester. Liksom proteinvikning sker glykosylering medan översättningen pågår.
Oligosackaridenheterna består av fjorton sockerrester. De överförs till asparagin med ett enzym som kallas oligosacaryltransferas, som ligger i membranet.
Medan proteinet finns i retikulumet avlägsnas tre glukosrester och en mannosrest. Dessa proteiner tas till Golgi-apparaten för vidare bearbetning..
Å andra sidan är vissa proteiner inte förankrade i plasmamembranet av en del av hydrofoba peptider. Däremot är de fästa vid vissa glykolipider som fungerar som ett förankringssystem och kallas glykosylfosfatidylinositol (förkortat GPI)..
Detta system är sammansatt i retikulummembranet och involverar bindning av GPI vid proteinets terminala kol..
Det endoplasmiska retikulumet spelar en avgörande roll i lipidsbiosyntesen; specifikt det släta endoplasmiska nätverket. Lipider är en oumbärlig komponent i plasmamembranen i celler.
Lipider är mycket hydrofoba molekyler, så de kan inte syntetiseras i vattenmiljöer. Därför sker dess syntes i samband med redan existerande membranformiga komponenter. Transporten av dessa lipider sker i vesiklar eller av transportproteiner.
Membranen i eukaryota celler består av tre typer av lipider: fosfolipider, glykolipider och kolesterol..
Fosfolipider härrör från glycerol och är de viktigaste strukturella beståndsdelarna. Dessa syntetiseras i regionen av retikulummembranet som pekar mot det cytosoliska ansiktet. Olika enzymer deltar i processen.
Membranet växer genom integrering av nya lipider. Tack vare förekomsten av enzymet flipas kan tillväxt förekomma i båda halvorna av membranet. Detta enzym är ansvarigt för att överföra lipider från ena sidan av dubbelskiktet till den andra..
Processerna för syntes av kolesterol och ceramider förekommer också i retikulum. Den senare reser till Golgi-apparaten för att producera glykolipider eller sfingomyelin..
Kalciummolekylen deltar som signaliserare i olika processer, oavsett om det är fusion eller förening av proteiner med andra proteiner eller med nukleinsyror..
Det inre av det endoplasmiska retikulumet har kalciumkoncentrationer på 100-800 uM. Kalciumkanaler och receptorer som frigör kalcium finns i retikulum. Kalciumfrisättning inträffar när fosfolipas C stimuleras genom aktivering av G-proteinkopplade receptorer (GPCR).
Vidare sker eliminering av fosfatidylinositol 4,5 bisfosfat i diacylglycerol och inositoltrifosfat; den senare ansvarar för frisättningen av kalcium.
Muskelceller har ett endoplasmatiskt retikulum som är specialiserat på bindning av kalciumjoner, kallat sarkoplasmatiskt retikulum. Det är involverat i processerna för muskelsammandragning och avslappning.


Ingen har kommenterat den här artikeln än.