De telofas det är det sista delningsstadiet av mitos och meios. Det är efter anafas och föregår cytoplasmisk uppdelning eller cytokinese. Det karakteristiska särdraget som skiljer och definierar det är bildandet av nya kärnor.
När det duplicerade DNA: t komprimerades (profas) migrerade de bundna systerkromatiderna till ekvatorn i cellen (metafas). När de alla var samlade där ställde de upp för att mobiliseras till delarna i den delande cellen under anafas..
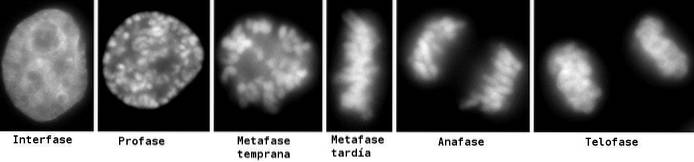
Slutligen, för att dela och ge upphov till två celler måste två kärnor först bildas för att skydda DNA. Detta är precis vad som händer under telofasen av mitos..
Inte för att något helt annat händer, mekaniskt sett, under telofaserna av meios I och meios II. Men materialen som tas emot som "kromosomer" är väldigt olika..
I telofas I mottar cellen i meios endast en uppsättning dubbla homologer vid varje pol. Det vill säga en enda uppsättning av kromosomkomplementet hos arten med varje kromosom bildad av två systerkromatider förenade med centromeren.
Vid telofasen av meios II migrerar systerkromatider poleward, och kärnor med ett haploida antal kromosomer bildas. I slutet av telofasen är kromosomerna inte längre synliga som kompakterade strukturer.
Artikelindex
I detta avsnitt kommer vi att överväga tre definierande aspekter av telofaser: början på bildandet av nukleoli, dekondensering av kromatin och uppkomsten av nya kärnhöljen..
I öppna mitoser bildas många små nukleoler, som när cykeln fortskrider, sammanfaller och bildar de nukleoler som är typiska för arten (som inte är många). Med händelser som utlöstes under metafas börjar den strukturella biogenesen av dessa organeller i telofas..
Detta är av stor betydelse eftersom i nukleolierna bland annat syntetiseras RNA som ingår i ribosomerna. I ribosomer utförs processen för translation av budbärar-RNA för att producera proteiner. Och varje cell, särskilt nya, behöver göra proteiner snabbt..
Genom att dela upp kommer därför varje ny cellprodukt från den divisionen att vara kompetent för översättningsprocessen och den autonoma existensen.
Å andra sidan är kromatinet som ärvs från anafas mycket komprimerat. Detta måste dekondenseras för att organisera det i kärnorna i bildning i öppna mitoser..
Rollen att kontrollera kromatindekondensering i en delande cell spelas av ett proteinkinas som kallas Aurora B. Detta enzym begränsar dekondensationsprocessen under anafas, vilket begränsar det till den sista fasen av division eller telofas. I själva verket är Aurora B proteinet som styr övergången från anafas till telofas..
Den andra viktiga aspekten av telofasen, och som definierar den, är bildandet av kärnhöljet. Låt oss komma ihåg att i öppna celldelningar försvinner kärnhöljet för att möjliggöra en fri mobilisering av det kondenserade kromatinet. Nu när kromosomerna har segregerat måste de grupperas i en ny kärna per cellpol.
För att generera en ny kärna måste kromatin interagera med de proteiner som kommer att bilda kärnkärna, eller lamininer. Lamininerna kommer i sin tur att fungera som en brygga för interaktionen med andra proteiner som gör det möjligt att bilda kärnkärna.
Detta kommer att separera kromatinet i eu- och heterokromatin, möjliggöra den inre organisationen av kärnan och hjälpa till att konsolidera det inre kärnmembranet..
Samtidigt kommer mikrotubulstrukturer härrörande från stamcellens endoplasmiska retikulum att migrera till kondensationszonen i telofaskromatin. De kommer att täcka över det i små fläckar och sedan smälta samman för att täcka det helt.
Detta är det externa kärnmembranet som är kontinuerligt med det endoplasmatiska retikulumet och med det inre kärnmembranet..
Alla ovanstående steg beskriver telosfasen av mitos i grunden. Vid varje cellpol bildas en kärna med modercellens kromosomala komplement..
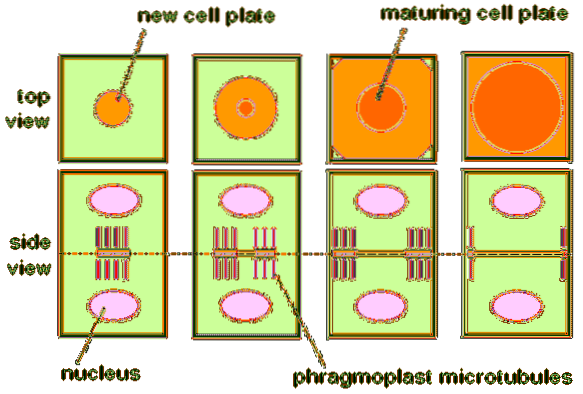
Men till skillnad från mitos hos djur bildas en unik struktur som kallas en fragmoplast under mitos i växtceller. Detta visas mellan de två framtida kärnorna i övergången mellan anafas och telofas..
Dess huvudroll i växt mitotisk uppdelning är att syntetisera cellplattan. Det vill säga fragmoplasten genererar platsen där de nya cellerna i växten kommer att dela sig när telofasen slutar..
I meiotiska telofaser sker det som redan har beskrivits, men med vissa skillnader. I telofas I bildas "kärnor" med en enda uppsättning homologa (duplicerade) kromosomer. Kärnor bildas i telofas II med ett haploida komplement av systerkromatider.
I många organismer förekommer inte kromatinkondensation vid telofas I, som övergår nästan omedelbart till meios II. I andra fall avkondenserar kromatinet, men kompakterar snabbt igen under profas II..
Kärnhöljet är vanligtvis kortlivat i telofas I, men permanent i II. Aurora B-proteinet kontrollerar segregeringen av homologa kromosomer under telofas I. Det deltar dock inte i segregeringen av systerkromatider under telofas II..
I alla fall av kärnuppdelning följs denna process av en uppdelning av cytoplasman, en process som kallas cytokinese. Cytokinese observeras både i slutet av telofas i mitos, och i slutet av telofas I och telofas II i meios.


Ingen har kommenterat den här artikeln än.