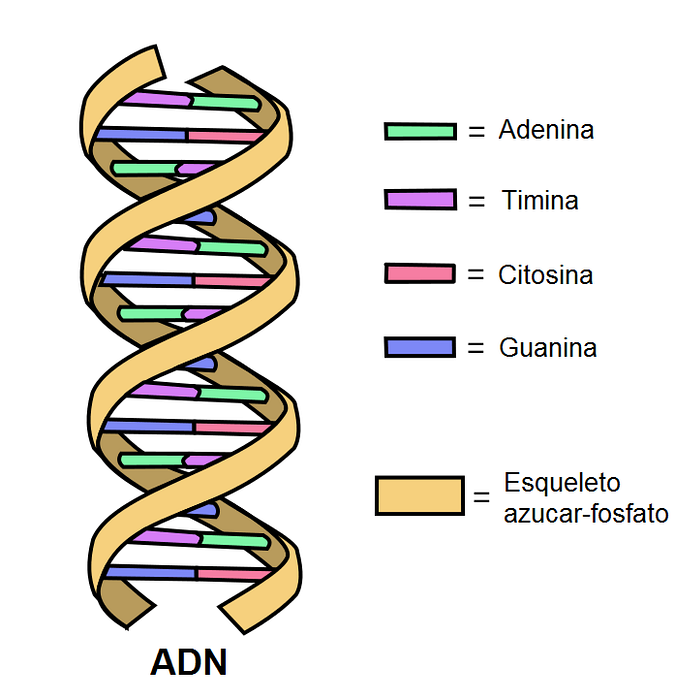
De DNA (deoxiribonukleinsyra) är den biomolekyl som innehåller all information som krävs för att generera en organism och bibehålla dess funktion. Den består av enheter som kallas nukleotider, består av en fosfatgrupp, en sockermolekyl med fem kol och en kvävehaltig bas..
Det finns fyra kvävebaser: adenin (A), cytosin (C), guanin (G) och tymin (T). Adenin parar alltid med tymin och guanin med cytosin. Meddelandet i DNA-strängen omvandlas till ett budbärar-RNA och detta deltar i syntesen av proteiner.

DNA är en extremt stabil molekyl, negativt laddad vid fysiologiskt pH, som associeras med positiva proteiner (histoner) för att effektivt komprimera i kärnan i eukaryota celler. En lång DNA-kedja bildar tillsammans med olika associerade proteiner en kromosom.
Artikelindex
1953 lyckades amerikanen James Watson och brittiska Francis Crick belysa DNA: s tredimensionella struktur tack vare det kristallografiska arbetet som utfördes av Rosalind Franklin och Maurice Wilkins. De baserade också sina slutsatser på andra författares arbete.
När DNA exponeras för röntgen bildas ett diffraktionsmönster som kan användas för att sluta strukturen av molekylen: en helix av två antiparallella kedjor som roterar till höger, där båda kedjorna förenas av vätebindningar mellan baserna. . Det erhållna mönstret var följande:

Strukturen kan antas enligt Braggs diffraktionslagar: när ett objekt placeras mitt i en röntgenstråle reflekteras det eftersom objektets elektroner interagerar med strålen..
Den 25 april 1953 publicerades resultaten av Watson och Crick i den prestigefyllda tidningen Natur, i en artikel på endast två sidor med titeln “Molekylär struktur av nukleinsyror”, Vilket skulle helt revolutionera området för biologi.
Tack vare denna upptäckt fick forskarna Nobelpriset i medicin 1962, med undantag av Franklin som dog före förlossningen. För närvarande är denna upptäckt en av de stora exponenterna för framgången för den vetenskapliga metoden att förvärva ny kunskap.
DNA-molekylen består av nukleotider, enheter som består av ett socker med fem kol som är fäst vid en fosfatgrupp och en kvävehaltig bas. Den typ av socker som finns i DNA är av deoxiribostyp och därav namnet deoxiribonukleinsyra..
För att bilda kedjan är nukleotiderna kovalent bundna av en fosfodiester-bindning genom en 3'-hydroxylgrupp (-OH) från ett socker och 5'-fosafofo av nästa nukleotid.
Nukleotider bör inte förväxlas med nukleosider. Den senare avser den del av nukleotiden som endast bildas av pentos (socker) och den kvävehaltiga basen.
DNA består av fyra typer av kvävebaser: adenin (A), cytosin (C), guanin (G) och tymin (T).
Kvävebaser klassificeras i två kategorier: puriner och pyrimidiner. Den första gruppen består av en ring med fem atomer fäst vid en annan ring av sex, medan pyrimidinerna består av en enda ring.
Av de nämnda baserna härstammar adenin och guanin från puriner. Däremot tillhör gruppen pyrimidiner tymin, cytosin och uracil (närvarande i RNA-molekylen).
En DNA-molekyl består av två kedjor av nukleotider. Denna "kedja" är känd som en DNA-sträng..
De två trådarna är länkade med vätebindningar mellan de komplementära baserna. Kvävebaser är kovalent kopplade till en ryggrad av socker och fosfater.
Varje nukleotid lokaliserad på en sträng kan kopplas till en annan specifik nukleotid på den andra strängen för att bilda den välkända dubbla spiralen. För att skapa en effektiv struktur kopplas A alltid med T med hjälp av två vätebindningar och G med C med tre broar..
Om vi studerar proportionerna av kvävebaser i DNA, kommer vi att upptäcka att mängden A är identisk med mängden T och densamma med G och C. Detta mönster är känt som Chargaffs lag.
Denna parning är energiskt gynnsam, eftersom den gör det möjligt att bevara en liknande bredd genom hela strukturen och bibehålla ett liknande avstånd längs socker-fosfat-ryggraden. Observera att en bas av en ring passar ihop med en av en ring.
Det föreslås att den dubbla spiralen består av 10,4 nukleotider per varv, åtskilda av ett centrum-till-centrum-avstånd på 3,4 nanometer. Valsningsprocessen ger upphov till bildandet av spår i strukturen, vilket kan observera ett större och ett mindre spår.
Spåren uppstår på grund av att glykosidbindningarna i basparen inte är motsatta varandra med avseende på deras diameter. Pyrimidin O-2 och purin N-3 finns i mindre spåret, medan huvudspåret ligger i motsatt region..
Om vi använder en stege-analogi, består stigarna av paren kompletterande baser till varandra, medan skelettet motsvarar de två greppskenorna..
Ändarna på DNA-molekylen är inte desamma, varför vi talar om en ”polaritet”. En av dess ändar, 3 ', bär en -OH-grupp, medan 5'-änden har den fria fosfatgruppen.
De två trådarna är placerade antiparallella, vilket innebär att de ligger mittemot sina polariteter, enligt följande:
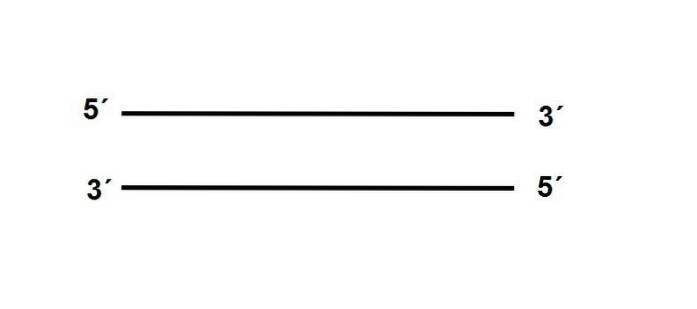
Dessutom måste sekvensen för en av strängarna vara komplementär till sin partner, om den är en position finns A, i den antiparallella strängen måste det finnas en T.
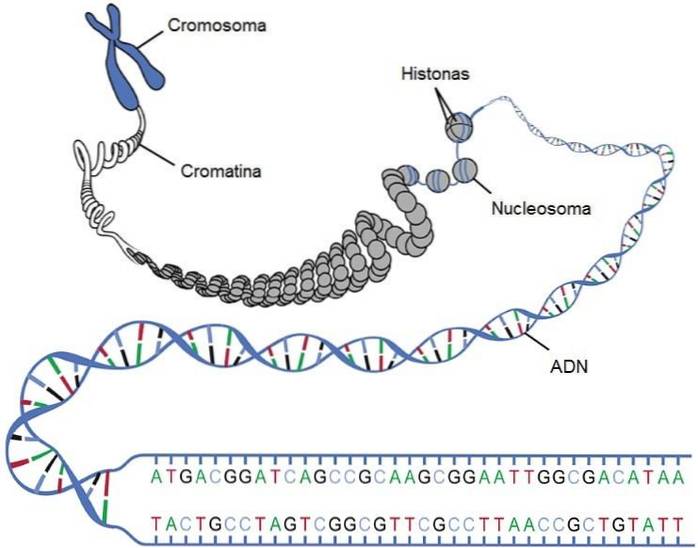
I varje mänsklig cell finns cirka två meter DNA som måste förpackas effektivt.
Strängen måste komprimeras så att den kan ingå i en mikroskopisk kärna med en diameter av 6 mikrometer som endast upptar 10% av cellvolymen. Detta är möjligt tack vare följande komprimeringsnivåer:
I eukaryoter finns proteiner som kallas histoner, som har förmågan att binda till DNA-molekylen, vilket är den första komprimeringsnivån av strängen. Histoner har positiva laddningar för att kunna interagera med de negativa laddningarna av DNA, tillhandahållna av fosfater.
Histoner är proteiner så viktiga för eukaryota organismer att de har varit praktiskt taget oförändrade under utvecklingsförloppet - kom ihåg att en låg grad av mutationer indikerar att det selektiva trycket på den molekylen är starkt. Histonskador kan leda till defekt DNA-komprimering.
Histoner kan modifieras biokemiskt och denna process ändrar komprimeringsnivån för det genetiska materialet.
När histoner är "hypoacetylerade" kondenseras kromatin mer, eftersom acetylerade former neutraliserar de positiva laddningarna av lysiner (positivt laddade aminosyror) i proteinet..
DNA-strängen rullar in i histonerna och de bildar strukturer som liknar pärlorna på ett pärlhalsband, kallat nukleosomer. Kärnan i denna struktur är två kopior av varje typ av histon: H2A, H2B, H3 och H4. Föreningen av de olika histonerna kallas "histonoktamer".
Oktamern omges av cirka 146 baspar, som kretsar mindre än två gånger. En human diploid cell innehåller cirka 6,4 x 109 nukleotider som är organiserade i 30 miljoner nukleosomer.
Organisation i nukleosomer gör att DNA kan komprimeras till mer än en tredjedel av sin ursprungliga längd.
I en process med extraktion av genetiskt material under fysiologiska förhållanden observeras att nukleosomer är ordnade i en 30-nanometerfiber.
Kromosomer är den funktionella enheten av ärftlighet, vars funktion är att bära en individs gener. En gen är ett segment av DNA som innehåller informationen för att syntetisera ett protein (eller en serie proteiner). Det finns dock också gener som kodar för regleringselement, såsom RNA.
Alla mänskliga celler (med undantag av könsceller och röda blodkroppar) har två kopior av varje kromosom, en ärvs från fadern och den andra från modern.
Kromosomer är strukturer som består av en lång linjär bit DNA associerad med proteinkomplexen som nämns ovan. Normalt i eukaryoter är allt genetiskt material som ingår i kärnan uppdelat i en serie kromosomer.
Prokaryoter är organismer som saknar en kärna. I dessa arter är det genetiska materialet mycket lindat tillsammans med alkaliska proteiner med låg molekylvikt. På detta sätt komprimeras DNA och ligger i en central region i bakterien..
Vissa författare kallar ofta denna struktur för "bakteriekromosom", även om den inte har samma egenskaper som en eukaryot kromosom..
Inte alla arter av organismer innehåller samma mängd DNA. Faktum är att detta värde är mycket varierande mellan arter och det finns inget samband mellan mängden DNA och organismens komplexitet. Denna motsägelse är känd som "C-värdeparadoxen".
Det logiska resonemanget skulle vara att intuitera att ju mer komplex organismen är, desto mer DNA har den. Detta är dock inte sant i naturen..
Till exempel lungfiskgenomet Protopterus aethiopicus Det är 132 pg i storlek (DNA kan kvantifieras i pikogram = pg) medan det mänskliga genomet bara väger 3,5 pg.
Man måste komma ihåg att inte allt DNA i en organism kodar för proteiner, en stor mängd av detta är relaterat till reglerande element och med olika typer av RNA.
Watson och Crick-modellen, härledd från röntgendiffraktionsmönstren, är känd som B-DNA-spiralen och är den ”traditionella” och mest kända modellen. Det finns dock två andra olika former, som kallas A-DNA och Z-DNA..
"A" -varianten vänder åt höger, precis som B-DNA, men är kortare och bredare. Denna form visas när den relativa luftfuktigheten minskar.
A-DNA roterar var 11: e baspar, huvudspåret är smalare och djupare än B-DNA. När det gäller mindre spår är detta mer ytligt och brett.
Den tredje varianten är Z-DNA. Det är den smalaste formen, bildad av en grupp hexanukleotider organiserade i en duplex av antiparallella kedjor. En av de mest framstående egenskaperna hos denna form är att den vänder till vänster, medan de andra två sätten gör det till höger..
Z-DNA uppträder när det finns korta sekvenser av pyrimidiner och puriner alternerande med varandra. Major sulcus är platt och minor är smal och djupare, jämfört med B-DNA.
Även om DNA-molekylen mestadels är i sin B-form under fysiologiska förhållanden, exponerar förekomsten av de två beskrivna varianterna det genetiska materialets flexibilitet och dynamik..
DNA-molekylen innehåller all information och instruktioner som är nödvändiga för att bygga en organism. Den kompletta uppsättningen genetisk information i organismer kallas genom.
Meddelandet kodas av det "biologiska alfabetet": de fyra baserna som nämnts tidigare, A, T, G och C.
Meddelandet kan leda till bildandet av olika typer av proteiner eller kod för något reglerande element. Processen genom vilken dessa databaser kan leverera ett meddelande förklaras nedan:
Meddelandet krypterat med de fyra bokstäverna A, T, G och C resulterar i en fenotyp (inte alla DNA-sekvenser kodar för proteiner). För att uppnå detta måste DNA replikera sig i varje celldelningsprocess..
DNA-replikering är halvkonservativ: en tråd tjänar som en mall för bildandet av den nya dottermolekylen. Olika enzymer katalyserar replikering, inklusive DNA-primas, DNA-helikas, DNA-ligas och topoisomeras..
Därefter måste meddelandet - skrivet i ett bassekvensspråk - överföras till en mellanliggande molekyl: RNA (ribonukleinsyra). Denna process kallas transkription..
För att transkription ska ske måste olika enzymer delta, inklusive RNA-polymeras.
Detta enzym ansvarar för att kopiera DNA-meddelandet och konvertera det till en budbärar-RNA-molekyl. Målet med transkription är med andra ord att få budbäraren.
Slutligen sker översättningen av meddelandet till budbärar-RNA-molekyler, tack vare ribosomerna.
Dessa strukturer tar budbärar-RNA och bildar tillsammans med översättningsmaskineriet det angivna proteinet..
Meddelandet läses i "tripletter" eller grupper med tre bokstäver som anger för en aminosyra - byggstenarna i proteiner. Det är möjligt att dechiffrera meddelandet från trillingarna eftersom den genetiska koden redan har avslöjats.
Översättningen börjar alltid med aminosyran metionin, som kodas av utgångstripletten: AUG. "U" representerar basuracil och är karakteristisk för RNA och ersätter tymin.
Till exempel, om budbärar-RNA har följande sekvens: AUG CCU CUU UUU UUA, översätts det till följande aminosyror: metionin, prolin, leucin, fenylalanin och fenylalanin. Observera att två tripletter - i detta fall UUU och UUA - kan koda för samma aminosyra: fenylalanin.
På grund av denna egenskap sägs det att den genetiska koden är degenererad, eftersom en aminosyra kodas av mer än en sekvens av tripletter, förutom aminosyran metionin, som dikterar translationens början..
Processen stoppas med specifika stopp- eller stopptripletter: UAA, UAG och UGA. De är kända under namnen på ockra, bärnsten och opal. När ribosomen upptäcker dem kan de inte längre lägga till fler aminosyror i kedjan.
Nukleinsyror är sura till sin natur och är lösliga i vatten (hydrofila). Bildningen av vätebindningar mellan fosfatgrupperna och hydroxylgrupperna av pentoser med vatten kan förekomma. Det är negativt laddat vid fysiologiskt pH.
DNA-lösningar är mycket viskösa på grund av den dubbla spiralens deformationsmotståndskapacitet, som är mycket stel. Viskositeten minskar om nukleinsyran är enkelsträngad.
De är mycket stabila molekyler. Logiskt sett måste denna egenskap vara oumbärlig i de strukturer som bär genetisk information. Jämfört med RNA är DNA mycket stabilare eftersom det saknar en hydroxylgrupp.
DNA kan värmedenatureras, vilket innebär att trådarna separeras när molekylen utsätts för höga temperaturer.
Mängden värme som måste appliceras beror på molekylens GC-procent, eftersom dessa baser är länkade av tre vätebindningar, vilket ökar motståndet mot separering..
När det gäller absorption av ljus har de en topp vid 260 nanometer, vilket ökar om nukleinsyran är enkelsträngad, eftersom nukleotidringarna är exponerade och dessa är ansvariga för absorptionen..
Enligt Lazcano et al. 1988 framträder DNA i övergångsfaser från RNA, som en av de viktigaste händelserna i livets historia.
Författarna föreslår tre steg: en första period där det fanns molekyler som liknade nukleinsyror, senare bestod genomet av RNA och som det sista steget uppstod dubbelbands-DNA-genomerna..
Vissa bevis stöder teorin om en primär värld baserad på RNA. För det första kan proteinsyntes förekomma i frånvaro av DNA, men inte när RNA saknas. Dessutom har RNA-molekyler med katalytiska egenskaper upptäckts..
När det gäller syntesen av deoxiribonukleotider (närvarande i DNA) kommer de alltid från minskningen av ribonukleotider (närvarande i RNA).
Den evolutionära innovationen av en DNA-molekyl måste ha krävt närvaron av enzymer som syntetiserar DNA-föregångare och deltar i den omvända transkriptionen av RNA.
Genom att studera nuvarande enzymer kan man dra slutsatsen att dessa proteiner har utvecklats flera gånger och att övergången från RNA till DNA är mer komplex än vad man tidigare trott, inklusive processer för överföring och förlust av gener och icke-ortologa ersättare..
DNA-sekvensering består av att belysa DNA-strängens sekvens i termer av de fyra baserna som komponerar den.
Kunskap om denna sekvens är av yttersta vikt i de biologiska vetenskaperna. Den kan användas för att skilja mellan två morfologiskt mycket liknande arter, för att upptäcka sjukdomar, patologier eller parasiter och har till och med en kriminalteknisk tillämpbarhet.
Sanger-sekvensering utvecklades på 1900-talet och är den traditionella tekniken för att klargöra en sekvens. Trots sin ålder är det en giltig metod och används ofta av forskare.
Metoden använder DNA-polymeras, ett mycket tillförlitligt enzym som replikerar DNA i celler, och syntetiserar en ny DNA-sträng med en befintlig som en guide. Enzymet kräver a först eller primer för att initiera syntes. Primern är en liten DNA-molekyl som är komplementär till molekylen som ska sekvenseras.
I reaktionen tillsätts nukleotider som kommer att införlivas i den nya DNA-strängen av enzymet.
Förutom de "traditionella" nukleotiderna innefattar metoden en serie dideoxynukleotider för var och en av baserna. De skiljer sig från standardnukleotider i två egenskaper: strukturellt tillåter de inte DNA-polymeras att lägga till fler nukleotider till dottersträngen och de har en annan fluorescerande markör för varje bas.
Resultatet är en mängd olika DNA-molekyler av olika längd, eftersom dideoxynukleotiderna införlivades slumpmässigt och stoppade replikationsprocessen i olika stadier..
Denna variation av molekyler kan separeras efter deras längd och nukleotidernas identitet avläses med hjälp av ljusemission från det fluorescerande märket..
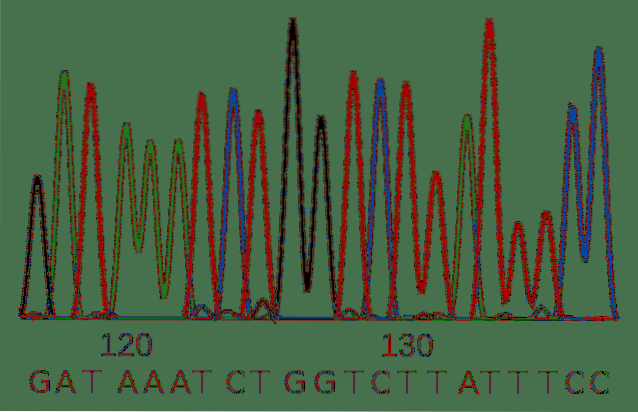
Sekvenseringstekniker som utvecklats under de senaste åren möjliggör massiv analys av miljontals prover samtidigt.
Bland de mest framstående metoderna är pyrosekvensering, sekvensering genom syntes, sekvensering genom ligering och nästa generations sekvensering av Ion Torrent..



Ingen har kommenterat den här artikeln än.