
De autofagi det är ett intracellulärt nedbrytningssystem som uppträder på ett konserverat sätt i lysosomerna i alla eukaryota celler (och jästvakuolerna). Ordet används vanligtvis för nedbrytningen av komponenterna i cytosolen eller "delarna" i cellen som är "föråldrade" eller som har slutat fungera ordentligt..
Termen autophagy myntades 1963 vid Rockefeller University av de Duve, som också observerade och beskrev processerna för cellulär endocytos. Bokstavligen betyder ordet autofagi "att konsumera sig själv", även om vissa författare beskriver det som en "självkannibalism".
Detta system skiljer sig från proteasomförmedlad nedbrytning genom att autofagi kan ta bort fullständiga intracellulära organeller och stora proteinkomplex eller aggregat på ett icke-selektivt sätt..
Trots denna icke-selektiva fagocytos har olika undersökningar visat att autofagi har många fysiologiska och patologiska konsekvenser. Eftersom det aktiveras under perioder av anpassning till svält, under utveckling, för eliminering av invaderande mikroorganismer, under programmerad celldöd, för eliminering av tumörer, presentation av antigener etc..
Artikelindex
Autophagy, som diskuterat, är en process som förmedlas av en cytoplasmatisk organell som kallas lysosomen..
Processen med "autofagi" börjar med inkapslingen av organellen som kommer att brytas ned av ett dubbelmembran och bilda en membrankropp som kallas autofagosomen. Autofagosommembranet smälter därefter med det lysosomala membranet eller med en sen endosom.
Var och en av dessa steg mellan sekvestrering, nedbrytning och frisättning av aminosyror eller andra komponenter för återvinning utövar olika funktioner i olika cellulära sammanhang, vilket gör autofagi till ett mycket multifunktionellt system..
Autophagy är en ganska kontrollerad process, eftersom endast de markerade cellulära komponenterna riktas mot denna nedbrytningsväg och markeringen sker i allmänhet under cellulära ombyggnadsprocesser..
Till exempel, när en levercell etablerar ett avgiftningsrespons som svar på fettlösliga läkemedel, förökas dess släta endoplasmatiska retikulum avsevärt, och när stimulansen som genereras av läkemedlet minskar, avlägsnas det överskott av jämnt endoplasmatiskt retikulum från det cytosoliska utrymmet genom autofagi..
En av de händelser som oftast utlöser autofagiska processer är svält..
Beroende på vilken organism som övervägs kan olika typer av väsentliga näringsämnen utlösa detta "återvinningssystem". I jäst, till exempel, även om bristen på kol i vissa aminosyror och nukleinsyror kan inducera autofagi, är bristen på kväve den mest effektiva stimulansen, vilket också gäller för växtceller.
Även om det inte har förstås helt, har celler speciella "sensorer" för att bestämma när ett näringsämne eller essentiell aminosyra är i mycket lågt tillstånd, och på så sätt utlösa hela återvinningsprocessen genom lysosomer.
Hos däggdjur deltar vissa hormoner i regleringen (positiv eller negativ) av autofagi i celler som tillhör vissa organ, såsom insulin, vissa tillväxtfaktorer eller interleukiner etc..
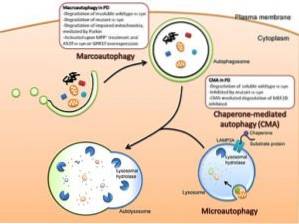
Det finns tre huvudtyper av autophagy bland eukaryoter: makro autophagy, micro autophagy och chaperone-medierad autophagy. Om inte annat anges avser termen autofagi makroautofagi..
Även om de tre typerna av autofagi är morfologiskt olika, slutar de alla i transporten av ämnen till lysosomer för nedbrytning och återvinning..
Detta är en typ av autofagi som beror på träningen de novo av fagocytiska vesiklar som kallas autofagosomer. Bildningen av dessa vesiklar är oberoende av bildandet av membran "knoppar", eftersom de bildas genom expansion.
I jäst börjar bildandet av autofagosomer vid ett visst ställe som kallas PAS, medan hos däggdjur förekommer många olika ställen i cytosolen, troligen kopplad till det endoplasmiska retikulumet genom strukturer som kallas "omegasomer".
Autofagosomernas storlek är mycket varierande och beror på organismen och typen av molekyl eller organell som är fagocyterad. Det kan variera från 0,4-0,9 mikrometer i diameter i jäst till 0,5-1,5 mikrometer hos däggdjur..
När autofagosomens och lysosomens membran smälter samman blandas innehållet i dessa och det är då matsmältningen av autofagins målsubstrat börjar. Denna organell är sedan känd som autolysosomen..
För vissa författare kan makroautofagi underklassificeras i sin tur till inducerad autofagi och autofag vid baslinjen. Inducerad makroautofagi används för att producera aminosyror efter en längre svältperiod.
Basal makroautofagi hänvisar till den konstitutiva mekanismen (som alltid är aktiv) som är väsentlig för omsättningen av de olika cytosoliska komponenterna och de intracellulära organellerna.
Denna typ av autofagi hänvisar till processen där det cytoplasmiska innehållet införs i lysosomen genom invaginationer som uppträder i organellens membran..
När de väl introducerats i lysosomen flyter blåsorna som produceras av dessa invaginationer fritt i lumen tills de lyseras och deras innehåll frigörs och bryts ned av specifika enzymer..
Denna typ av autofagi har endast rapporterats för däggdjursceller. Till skillnad från makro-autofagi och mikro-autofagi, där vissa cytosoliska delar är icke-specifikt fagocytos, är autofagi medierad av chaperoner ganska specifik, eftersom det beror på närvaron av särskilda pentapeptidsekvenser i substraten som kommer att fagocytoseras..
Vissa forskare har bestämt att detta pentapeptidmotiv är relaterat till KFERQ-sekvensen och att det finns i mer än 30% av cytosolproteiner.
Det kallas "chaperone-medierad" eftersom chaperonproteiner är ansvariga för att hålla detta konserverade motiv exponerat för att underlätta dess igenkänning och förhindra att proteinet fälls på det..
Proteiner med denna tagg translokeras till lysosomal lumen och där bryts de ned. Många av nedbrytningssubstraten är glykolytiska enzymer, transkriptionsfaktorer och deras hämmare, kalcium- eller lipidbindande proteiner, proteasom-underenheter och vissa proteiner involverade i vesikulär handel..
Liksom de andra två typerna av autophagy är chaperon-medierad autophagy en reglerad process på många nivåer, från etikettigenkänning till transport och nedbrytning av substrat i lysosomer..
En av huvudfunktionerna för den autofagiska processen är eliminering av åldrande eller "föråldrade" organeller, som är märkta med olika vägar för deras nedbrytning inom lysosomer..
Tack vare observationen av elektronmikrofotografier av lysosomer i däggdjursceller har närvaron av peroxisomer och mitokondrier detekterats i dem..
I en levercell är till exempel en mitokondrions genomsnittliga livslängd 10 dagar, varefter denna organell fagocyteras av lysosomer, där den bryts ned och dess komponenter återvinns för olika metaboliska ändamål..
Under förhållanden med låg näringskoncentration kan celler utlösa bildandet av autofagosomer för att selektivt "fånga" delar av cytosolen, såväl som de smälta metaboliterna i dessa autofagosomer kan hjälpa celler att överleva när yttre förhållanden är begränsade ur synvinkel. näringsmässig synvinkel.
Autophagy har viktiga funktioner i omstruktureringen av celler i differentieringsprocessen, eftersom det deltar i kassering av cytosoliska delar som inte krävs vid specifika tidpunkter..
Det har också viktiga konsekvenser för cellhälsan, eftersom det är en del av försvarsmekanismerna mot invaderande virus och bakterier..
Yoshinori Ohsumi, en Nobelprisbelönt japansk forskare inom fysiologi och medicin 2016, beskrev de molekylära mekanismerna för autofagi i jäst medan han studerade det metaboliska ödet för många proteiner och vakuolerna hos dessa encelliga organismer..
I sitt arbete identifierade Ohsumi inte bara proteinerna och vägarna som var inblandade i processen utan visade också hur autofagvägen regleras tack vare verkan av proteiner som kan "räkna" olika metaboliska tillstånd..
Deras arbete började med exakta mikroskopiska observationer av vakuoler under intensiva nedbrytningshändelser. Vakuoler betraktas som lagringsplatser för jäst ”skräp” och cellulärt skräp..
Genom att observera jäst med defekta mutantgenotyper för olika gener relaterade eller hypotetiskt relaterade till autofagi (känd som generna ATG) kunde denna forskare och hans medarbetare beskriva det autofagiska jästsystemet på genetisk nivå.
Därefter bestämde denna grupp forskare de viktigaste genetiska egenskaperna hos proteinerna som kodas av dessa gener och gav betydande bidrag till deras interaktion och bildandet av de komplex som var ansvariga för initiering och utförande av autofagi i jäst..
Tack vare arbetet med Yoshinori Ohsumi förstår vi idag bättre de molekylära aspekterna av autofagi, liksom dess viktiga konsekvenser för att cellerna och organen som utgör oss fungerar korrekt..


Ingen har kommenterat den här artikeln än.