De könsceller de är föregångarcellerna hos könscellerna hos de djur med sexuell reproduktion. De är embryonala föregångare som skiljer sig från flera somatiska cellinjer (soma = kropp) mycket tidigt i utvecklingen av de flesta arter.
Praktiskt taget alla organismer som reproducerar sexuellt uppstår genom fusion av två gametiska celler. Gameter är specialiserade celler som bär hälften av den genetiska informationen hos individen som producerar dem, fadern och modern (de är haploida celler).
Alla könsceller som produceras av ett djur bildas från en speciell celllinje som kallas groddlinjen, som utvecklas enligt en komplex uppsättning specifika signaler. Dessa celler representerar den huvudsakliga "överföringsvägen" för genomet och cytosoliska komponenter från en generation till en annan..
Könsceller är ansvariga för processerna för speciering och evolution, eftersom det är förändringarna som sker i dessa som överförs från en generation till en annan. Dessutom är dessa celler de som förmedlar överföring av ärftliga sjukdomar från föräldrar till sina barn, särskilt hos människor..
Artikelindex
Könsceller är "pluripotenta" eller "totipotenta" embryonala celler, det vill säga de kan differentieras till nästan vilken celltyp som helst under rätt förhållanden och signaler. Dessutom är de kompetenta celler för sin "självförnyelse", eftersom de är ansvariga för sin egen regenerering..
Dessa celler är de enda som kan producera könsceller, vilka är de celler som kan bilda en ny organism, en egenskap som de andra cellerna i ett embryo förlorar när de differentierar..
Vissa författare anser dem alltså som "stamceller" hos en art, eftersom de inte bildar organ utan nya individer. På samma sätt är dessa celler det huvudsakliga sättet att arter utvecklas och är medel för överföring av ärftliga sjukdomar, särskilt hos människor..
Könsceller producerar könsceller genom processer som kallas meios och gametogenes (oogenes och spermatogenes hos många djur), som är karakteristiska och unika för denna cellgrupp..
Könsceller skiljer sig tidigt från andra somatiska cellinjer under embryonal utveckling.

I många arter, såsom fruktflugan D. melanogaster, Dessa celler bildas av urceller i blastula som har "ärvt" en cytosolisk determinant känd som "grodplasma" eller "grodplasma", det vill säga mycket speciella blastomerer..
Nämnda bakterieplasma innehåller strukturella element och en del budbärar-RNA och under oogenes och befruktning genomgår det olika cytosoliska rörelser, för att senare bilda urcellkluster i blastula-steget, vilket kommer att ge upphov till primordiala könsceller.
Blastomererna som har "grodplasmen" delar sig asymmetriskt och arver grodplasmen till en enda dottercell. När embryot når gastrulastadiet, börjar motsvarande delning av dessa celler och populationen av urceller i groddlinjen expanderar..
Hos däggdjur som människa har å andra sidan inte involvering av en ”bakterieplasma” rapporterats under processen för ursprunglig bakteriecellbildning, utan snarare bestäms specifikationen av denna linje av cell-cell-interaktioner..
Urkimceller, under de tidiga stadierna av embryogenes, är belägna i ett slags extra-embryonalt fack och hos människor inträffar detta runt den tredje utvecklingsveckan.
När den ursprungliga cellinjen har definierats migrerar de mot kvinnliga eller manliga könsorganen, där oogenes- eller spermatogenesprocesserna aktiveras..
Samspelet mellan urcellerna och de somatiska cellerna i könsorganen, förutom närvaron av könskromosomerna och andra moderfaktorer, är det som definierar bestämningen av kön i groddlinjen, även om denna process kan variera avsevärt mellan en art och annan.
För att den initiala "separationen" av somatiska celler och könsceller ska äga rum är det första som inträffar ett differentiellt uttryck av gener, eftersom i groddlinjen undertrycks de somatiska linjernas karakteristiska gener för att starta "programmet Germ cellgenetik.
Under deras bildningsprocess migrerar dessa celler också från varifrån de kommer till den specifika platsen där könsorganen kommer att bildas, vilka är de könsproducerande vävnaderna hos vuxna..
Cellmigrering uppnås också genom aktivering av en hel migrerande "maskin" och olika "vägledande" mekanismer, som har att göra med flera genetiska och epigenetiska faktorer (som inte har något att göra med modifiering av nukleotidsekvensen).
Urkimceller, de som ger upphov till de "sanna" könscellerna, bildar långt ifrån var de utvecklas och måste, för att etablera sig, migrera till de platser där äggstockarna och testiklarna finns, vilka är kvinnliga och manliga könsorganen..
Urkimceller ses initialt under gastrulering som ett kluster av celler vid basen av allantois, vilket är ett extraembryonalt membran som bildas som ett utflöde från embryonets primära matsmältningskanal..
I detta skede förvärvar urceller en polariserad morfologi och vissa experiment har visat att de förlänger långa processer när de mobiliseras..
Senare blir dessa uppenbara i tarmen och kommer sedan ut från tarmryggen och migrerar i sidled och koloniserar könsryggarna..
När primordialcellerna rör sig från tarmen till den omgivande bindväven förlängs den senare och bildar tarmmesenteriet (vävnaden som täcker tunntarmen och förenar den med bukväggen), en process som inträffar medan celler dyker upp genom väggen av tarmen.
Ankomsten av föregångarcellerna mot gonadvävnaden styrs av de somatiska cellerna i nämnda strukturer, vilka uppenbarligen utövar en "kemoattraktiv" effekt på de förstnämnda..
Det har experimentellt visats att uttrycket av en gen känd som fragilis har mycket att göra med utvecklingen av rörlighet i urkimceller.
Denna gen är involverad i cellcellsadhesionsprocesser och i kontrollen av cellcykeln, så det misstänks att regleringen av vidhäftningsprocesser kan vara avgörande för initieringen av migrationsprocessen.
Vissa författare anser emellertid att migrationsvägen för dessa celler kontrolleras på miljönivå istället för att vara en autonom process..
På väg till könsorganen och en gång i dem multiplicerar dessa celler med mitos och genererar flera kloner som lyckas öka antalet individer i cellpopulationen..
När de ursprungliga könscellerna når sina slutliga vävnader utvecklas de till kvinnliga könsceller eller manliga könsceller, beroende på fallet och de endogena och exogena signalerna de får..
Kvinnliga könsorgan är äggstockarna och de manliga könsorganen är testiklarna. En gång i dessa vävnader multiplicerar urcellerna med hög hastighet, men mönstren för denna mitotiska spridning skiljer sig mellan de två..
Det finns alltså två typer av könsceller som kallas oogonia och spermatogonia..
Oogonia är mitotiskt aktiva celler. De delar sig intensivt under embryonal utveckling, specifikt från andra till femte graviditetsmånaden hos människor, med vilka upp till mer än 7 miljoner av dessa celler bildas, även om vissa degenererar naturligt.
Dessa celler delar sig inte genom mitos igen i postnatala stadier, utan differentieras i stället periodiskt. Under de sista stadierna av fosterutvecklingen börjar de dock dela sig med meios, en process som förblir i "arrest" fram till början av puberteten..
Spridningen av spermatogonia är något annorlunda än den för oogonia, för trots att de börjar bildas och föröka sig i embryonala testiklar, behåller de sin förmåga att dela upp i praktiskt taget hela det postnatala livet..
Testiklarnas seminiferous kanaler är avgränsade internt med spermatogonia, och några av populationerna som består av dessa celler delar sig med mitos. I början av puberteten börjar grupper av spermatogonia (primära spermatocyter) dela sig med meios för att bilda sekundära spermatocyter som ger upphov till haploida spermatider..
Könsceller är "fabrikerna" där "fordon" för överföring av information från en generation till nästa produceras. På samma sätt är dessa celler av stor betydelse för evolutionära processer, eftersom nästan alla modifieringar de genomgår kommer att präglas på avkomman..
Vi kan säga att DNA i alla celler i en organism är benägen för mutationer och även om mutationer i somatiska celler är viktiga i samband med många sjukdomar och andra tillstånd, sträcker de sig inte alltid längre än organismens livslängd. bär dem.
Germline-mutationer, å andra sidan, bidrar direkt till evolutionära genetiska processer, eftersom dessa förändringar kan överföras från en generation till nästa genom könsceller och zygoter..
Av dessa skäl kan mutationer i könsceller endast vara synliga i avkomman och detta beror mycket ofta på homozygositet eller heterozygositet hos de drabbade generna hos varje förälder..
Orsakerna till könslinjemutationer är många, eftersom de kan uppstå som svar på endogena eller exogena signaler. Vissa av dessa mutationer producerar sjukdomar som kan ärvas genom moderlinjen eller faderns linje, beroende på fallet..
Den okontrollerade delningen av celler i nästan vilken vävnad som helst i människokroppen, liksom hos andra djur, kan resultera i bildandet av tumörer, som kan vara godartade eller maligna..
De som härrör från könsceller kallas vanligtvis neoplasmer och kan vara:
- Groddar
- Teratomas
- Embryonal karcinom
- Endodermala sinustumörer
- Koriokarcinom
Dessa tumörer kan förekomma regelbundet i de inre regionerna i könsorganen, även om de också kan relateras till spridning eller avvikande migrering av primära könsceller, vilket innebär att de kan förekomma i olika delar av kroppen.
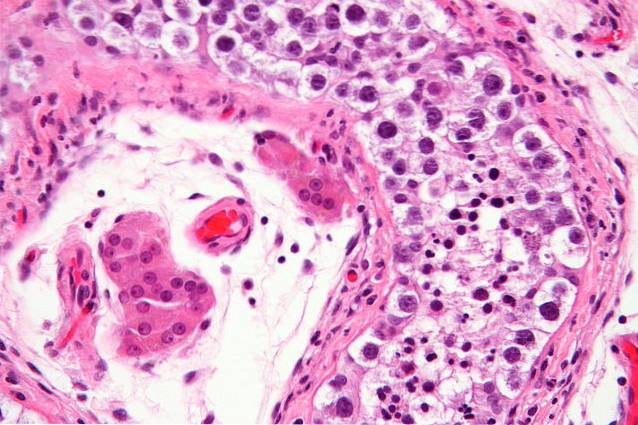
Tumörer associerade med den ursprungliga könscellinjen är kända som germinom, medan embryonala karcinom är de som härrör från embryonala "stamceller" eller härledda celler..
Vanligtvis elimineras de ursprungliga könscellerna som bildas i extragonadala platser, men teratomer är avvikande tillväxt av extragonadala bakterieceller som har lyckats överleva, och som består av slumpmässiga blandningar av differentierade vävnader såsom brosk, hud, hår eller tänder..
Endodermala sinustumörer är de som bildas från celler härrörande från extraembryonala vävnader och som är differentierade som bildar endodermal äggula. Om tvärtom tvärtom bildas i det trofoblastiska skiktet kallas det koriokarcinom.
Könsceller tumörer i äggstockarna står för cirka 20% av alla äggstockstumörer, är vanliga hos flickor och unga vuxna upp till cirka 20 år och är nästan alltid teratomer av malign karaktär..
Bland dessa har dysgerminom särskiljts, vilka är solida och köttiga tumörer som har ett mjukt lock, som består av aggregat av celler med ett polygonal utseende, med framstående plasmamembran och ett stort antal cytosoliska granuler..



Ingen har kommenterat den här artikeln än.