De zinkfingrar (ZF) är strukturmotiv som finns i ett stort antal eukaryota proteiner. De tillhör gruppen metalloproteiner, eftersom de kan binda den zinkmetalljon, som de behöver för att fungera. Mer än 1 500 ZF-domäner förutspås förekomma i cirka 1000 olika proteiner hos människor..
Uttrycket zinkfinger eller "zinkfinger" myntades först 1985 av Miller, McLachlan och Klug, medan man studerade i detalj de små DNA-bindande domänerna för transkriptionsfaktorn TFIIIA av Xenopus laevis, beskrivs av andra författare några år tidigare.
Proteiner med ZF-motiv är bland de vanligaste i genomet av eukaryota organismer och deltar i en mängd viktiga cellulära processer, inklusive gentranskription, proteinöverföring, metabolism, vikning och sammansättning av andra proteiner och lipider., Programmerad celldöd, bland andra.
Artikelindex
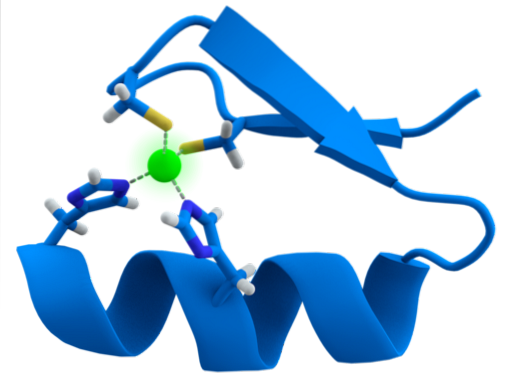
ZF-motivens struktur är extremt bevarad. Vanligtvis har dessa upprepade regioner 30 till 60 aminosyror, vars sekundära struktur återfinns som två antiparallella beta-ark som bildar en hårnål och en alfa-helix, som betecknas som ββ.
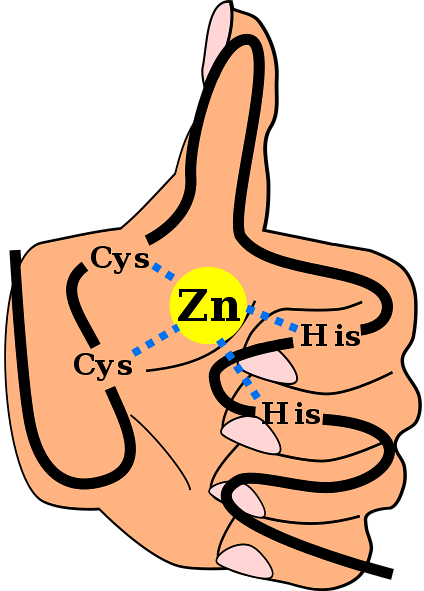
Nämnda sekundära struktur stabiliseras genom hydrofoba interaktioner och genom koordinering av en zinkatom som ges av två cystein- och två histidinrester (CystvåHanstvå). Det finns dock ZF som kan samordna mer än en zinkatom och andra där Cys och hans rester varierar..
ZF: erna kan hittas upprepade i satser, linjärt konfigurerade i samma protein. Alla har liknande strukturer, men kan skilja sig kemiskt från varandra genom variationer av aminosyrarester som är viktiga för att uppfylla sina funktioner.
Ett vanligt inslag bland ZF är deras förmåga att känna igen DNA- eller RNA-molekyler av olika längd, varför de ursprungligen endast ansågs som transkriptionsfaktorer.
I allmänhet är igenkänning av 3bp-regioner i DNA och uppnås när proteinet med ZF-domän presenterar alfa-helix till DNA-molekylens huvudspår.
Det finns olika ZF-motiv som skiljer sig från varandra genom sin natur och de olika rumsliga konfigurationerna som uppnås genom koordineringsbindningarna med zinkatomen. En av klassificeringarna är som följer:
Detta är ett vanligt förekommande motiv i ZF. De flesta C-motivtvåHtvå De är specifika för interaktion med DNA och RNA, men de har observerats delta i protein-protein-interaktioner. De har mellan 25 och 30 aminosyrarester och finns inom den största familjen av reglerande proteiner i däggdjursceller.
De interagerar med RNA och några andra proteiner. De observeras huvudsakligen som en del av vissa retroviruscapsidproteiner, vilket hjälper till med förpackningen av viralt RNA strax efter replikering..
Proteiner med detta motiv är enzymer som ansvarar för DNA-replikering och transkription. Ett bra exempel på dessa kan vara de grova enzymerna för fag T4 och T7.
Denna ZF-familj innefattar transkriptionsfaktorer som reglerar uttrycket av viktiga gener i många vävnader under cellutveckling. Faktorerna GATA-2 och 3 är till exempel involverade i hematopoies.
Dessa domäner är typiska för jäst, speciellt GAL4-proteinet, som aktiverar transkriptionen av gener som är involverade i användningen av galaktos och melibios..
Dessa specifika strukturer har två undertyper av ZF-domäner (C3HC4 och C3HtvåC3) och finns i många animaliska och växtproteiner.
De finns i proteiner som RAD5, involverade i DNA-reparation i eukaryota organismer. De finns också i RAG1, vilket är viktigt för omkonfigurering av immunglobuliner.
Denna ZF-domän är mycket konserverad i integraserna av retrovirus och retrotransposoner; genom att binda till målproteinet orsakar det en konformationsförändring i det.
Proteiner med ZF-domäner tjänar olika syften: de finns i ribosomala proteiner eller i transkriptionsadaptrar. De har också detekterats som en integrerad del av strukturen av jäst-RNA-polymeras II..
De verkar vara inblandade i intracellulär zinkhomeostas och i regleringen av apoptos eller programmerad celldöd. Dessutom finns det några proteiner med ZF som fungerar som chaperoner för vikning eller transport av andra proteiner..
Bindning till lipider och en grundläggande roll i protein-protein-interaktioner är också framstående funktioner för ZF-domänerna i vissa proteiner..
Genom åren har den strukturella och funktionella förståelsen av ZF-domänerna möjliggjort stora vetenskapliga framsteg som innebär att deras egenskaper används för biotekniska ändamål..
Eftersom vissa proteiner med ZF har hög specificitet för vissa DNA-domäner, investeras för närvarande mycket ansträngningar i utformningen av specifika ZF, vilket kan ge värdefulla framsteg inom genterapi hos människor..
Intressanta biotekniska tillämpningar uppstår också från designen av proteiner med genetiskt modifierade ZF. Beroende på det önskade syftet kan vissa av dessa modifieras genom tillsats av "polyzink" -pingerpeptider, vilka kan känna igen praktiskt taget vilken DNA-sekvens som helst med hög affinitet och specificitet..
Nukleasmodifierad genomredigering är en av de mest lovande applikationerna idag. Denna typ av redigering ger möjlighet att genomföra studier om genetisk funktion direkt i det intressanta modellsystemet..
Genteknik med modifierade ZF-nukleaser har fångat forskarnas uppmärksamhet inom området genetisk förbättring av sorter av växter av agronomisk betydelse. Dessa nukleaser har använts för att korrigera en endogen gen som producerar herbicidresistenta former i tobaksplantor..
Nukleaser med ZF har också använts för tillsats av gener i däggdjursceller. Proteinerna i fråga användes för att generera en uppsättning isogena musceller med en serie alleler definierade för en endogen gen..
En sådan process har en direkt tillämpning vid märkning och skapande av nya allelformer för att studera förhållandena mellan struktur och funktion under naturliga uttrycksförhållanden och i isogena miljöer..



Ingen har kommenterat den här artikeln än.