De erytroblaster de är föregångarceller av ryggradsdjur erytrocyter. Minskningen av syrekoncentrationen i vävnaderna kommer att främja celldifferentieringshändelser i dessa celler som ger upphov till mogna erytrocyter. Uppsättningen av alla dessa händelser är känd som erytropoies.
Under erytropoies ökar hemoglobinsyntesen. Ett rikligt protein i erytrocyter som förmedlar syretillförsel till vävnader och avgiftning av koldioxid från vävnader, en celltoxisk avfallsprodukt från cellulär andning.
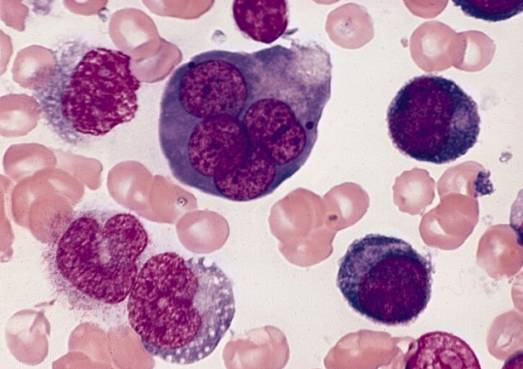
Fel i differentieringsprocessen av erytroblaster ger upphov till en uppsättning blodpatologier som tillsammans kallas megaloblastiska anemier.
Artikelindex
De har en karaktäristisk morfologi som liknar bikonkava skivor och deras huvudsakliga funktion är att utföra transporten av syre (O2) till kroppens olika vävnader, samtidigt som det avgiftar dem från koldioxiden (CO2) som produceras under cellulär andning..
Detta utbyte av CO2 mot O2 är möjligt eftersom dessa celler innehåller stora mängder av ett karakteristiskt rött protein som kallas hemoglobin, som kan interagera med båda kemiska arterna genom en hemgrupp som finns i deras struktur..
En särdrag hos dessa celler hos däggdjur med avseende på resten av ryggradsdjur är bristen på kärnor och cytoplasmatiska organeller. Under de första faserna av produktionen i de tidiga stadierna av embryonal utveckling har det emellertid observerats att de cellulära föregångarna från vilka de härstammar utgör en övergående kärna.
Det senare är inte förvånande med tanke på att de tidiga stadierna av embryoutveckling vanligtvis är lika i alla ryggradsdjur, vilket bara skiljer sig åt de stadier som äventyrar större differentiering..
Erytroblaster är celler som kommer att ge upphov till mogna erytrocyter efter att ha genomgått på varandra följande händelser av celldifferentiering..
Dessa föregångarceller härstammar från en vanlig myeloid stamfader i ryggradsdjurens benmärg som kärnformade celler, försedda med kärnor och cellulära organeller..
Förändringar i innehållet i dess cytoplasma och i omorganiseringen av cytoskelettet kommer att kulminera i genereringen av erytrocyter redo för cirkulation. Dessa förändringar svarar på miljöstimuli som indikerar minskningen av syre i vävnaderna och därför ett behov av produktion av erytrocyter..
Erytropoies är den term som används för att definiera den process genom vilken produktion och utveckling av röda blodkroppar sker, nödvändigt för att upprätthålla syretillförseln till de olika organen och vävnaderna..
Denna process regleras fint av verkan av erytropoietin (EPO), ett renalt synteshormon som i sin tur moduleras av de syrekoncentrationer som finns i vävnaderna..
Låga syrehalter i vävnaden inducerar EPO-syntes genom hypoxiinducerbar transkriptionsfaktor (HIF-1), vilket stimulerar erytrocytproliferation genom bindning till EpoR-receptorer, närvarande på erytrocytprekursorceller.
Hos däggdjur utförs erytropoies i två steg som kallas primitiv erytropoies och definitiv erytropoies..
Den första inträffar i äggulansäcken under embryonal utveckling, vilket ger upphov till stora kärnformade erytroblaster, medan den andra förekommer i fostrets lever och fortsätter i benmärgen efter den andra graviditetsmånaden och genererar mindre enuklerade erytrocyter..
Andra proteiner såsom den antipoptotiska cytokinen Bcl-X, vars transkription regleras av transkriptionsfaktorn GATA-1, påverkar också erytropoiesprocessen positivt. Dessutom är leveransen av järn, vitamin B12 och folsyra också nödvändig.
I processen för definitiv erytropoies bildas erytrocyter i benmärgen från en odifferentierad stamcell eller vanlig myeloid stamfader som kan ge upphov till andra celler såsom granulocyter, monocyter och trombocyter..
Denna cell måste ta emot lämpliga extracellulära signaler för att äventyra dess differentiering mot erytroids härstamning..
När detta åtagande har förvärvats börjar en sekvens av differentieringshändelser som börjar med bildandet av pronormoblasten, även känd som proerythroblast. En stor föregångarcell med erytroblast med en kärna.
Därefter kommer proerythroblast att uppleva en progressiv minskning i kärncellvolym åtföljd av en ökning av hemoglobinsyntes. Alla dessa förändringar sker långsamt när denna cell passerar genom olika cellsteg: basofil erytroblast eller normoblast, polykromatisk erytroblast och ortokromatisk erytroblast.
Processen avslutas med den totala förlusten av kärnan, såväl som av de organeller som finns i den ortokromatiska erytroblasten, med ursprung i en mogen erytrocyt..
För att äntligen nå detta måste den senare passera genom retikulocytstadiet, en enucleated cell som fortfarande innehåller organeller och ribosomer i dess cytoplasma. Fullständigt avlägsnande av kärnan och organellerna utförs genom exocytos.
Mogna erytrocyter lämnar benmärgen in i blodomloppet där de förblir cirkulerande i cirka 120 dagar innan de slukas av makrofager. Därför är erytropoies en process som sker kontinuerligt under hela organismen..
När erytoblaster utvecklas mot fullständig differentiering till en mogen erytrocyt genomgår de flera förändringar i sitt cytoskelett, liksom i uttrycket av celladhesionsproteiner..
Actin-mikrofilament depolymeriserar och ett nytt spektrinbaserat cytoskelett monteras. Spectrin är ett perifert membranprotein beläget på det cytoplasmiska ansiktet som interagerar med ankyrin, ett protein som förmedlar bindningen av cytoskelettet med Band 3-transmembranproteinet.
Dessa förändringar i cytoskelettet och i uttrycket av Epo-receptorer, liksom de mekanismer som modulerar dem, är kritiska för erytroid mognad..
Detta beror på det faktum att de förmedlar etableringen av interaktioner mellan erytroblaster och celler som finns i benmärgsmikromiljön, vilket underlättar överföringen av nödvändiga signaler för att påbörja och avsluta differentiering..
När differentieringen är klar inträffar nya förändringar som gynnar förlusten av cellernas vidhäftning till märgen och deras frisättning i blodomloppet där de kommer att utföra sin funktion..
Fel vid differentiering av erytroblaster i benmärgen ger upphov till blodpatologier, såsom megaloblastiska anemier. Dessa härrör från brister i leveransen av vitamin B12 och folat som är nödvändiga för att främja erytroblastdifferentiering..
Uttrycket megaloblastisk hänvisar till den stora storlek som erytroblaster och till och med erytrocyter når som en produkt av ineffektiv erytropoies som kännetecknas av defekt DNA-syntes.



Ingen har kommenterat den här artikeln än.