De helikas avser en grupp enzymer av den proteinhydrolytiska typen som är mycket viktiga för alla levande organismer; de kallas också motorproteiner. Dessa rör sig genom cellcytoplasman och omvandlar kemisk energi till mekaniskt arbete genom ATP-hydrolys..
Dess viktigaste funktion är att bryta vätebindningarna mellan de kvävehaltiga baserna av nukleinsyror, vilket möjliggör replikering av dem. Det är viktigt att betona att helikaser är praktiskt taget allestädes närvarande, eftersom de finns i virus, bakterier och i eukaryota organismer..
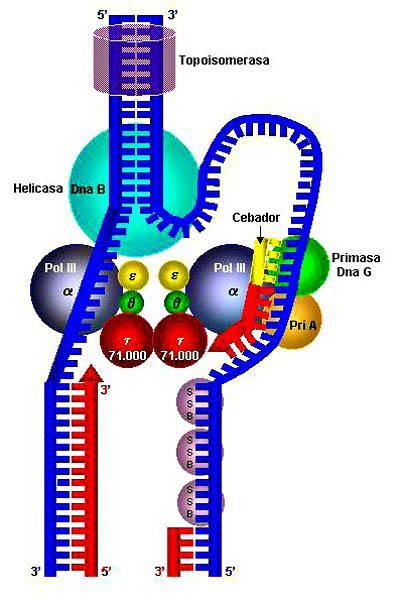
Det första av dessa proteiner eller enzymer upptäcktes 1976 i bakterier Escherichia coli; två år senare upptäcktes den första helikasen i en eukaryot organism, i liljväxter.
För närvarande har helikasproteiner karaktäriserats i alla naturliga riken inklusive virus, vilket innebär att en stor kunskap har genererats om dessa hydrolytiska enzymer, deras funktioner i organismer och deras mekanistiska roll..
Artikelindex
Hellikaser är biologiska eller naturliga makromolekyler som påskyndar kemiska reaktioner (enzymer). De kännetecknas huvudsakligen av att separera kemiska komplex av adenosintrifosfat (ATP) genom hydrolys.
Dessa enzymer använder ATP för att binda och modernisera komplex av deoxiribonukleinsyror (DNA) och ribonukleinsyror (RNA)..
Det finns minst två typer av helikaser: DNA och RNA.
DNA-helikaser verkar i DNA-replikering och kännetecknas av att separera dubbelsträngat DNA i enstaka strängar.
Dessa enzymer verkar i de metaboliska processerna för ribonukleinsyra (RNA) och vid multiplikation, reproduktion eller ribosomal biogenes.
RNA-helikas är också nyckeln till pre-splitsningsprocessen för budbärar-RNA (mRNA) och initieringen av proteinsyntes efter transkription av DNA till RNA i cellkärnan..
Dessa enzymer kan differentieras enligt deras aminosyrasekvenseringshomologi med kärnaminosyra-ATPas-domänen, eller genom delade sekvenseringsmotiv. Enligt klassificeringen är dessa grupperade i 6 superfamiljer (SF 1-6):
Enzymerna i denna superfamilj har en 3'-5 'eller 5'-3' translokationspolaritet och bildar inte ringstrukturer..
Det är känt som den största gruppen helikaser och består huvudsakligen av RNA-helikaser. De har en translokationspolaritet i allmänhet 3'-5 'med mycket få undantag.
De har nio motiv (från engelska motiv, som översätts som "återkommande element") av högt konserverade aminosyrasekvenser och, som SF1, bildar inte ringstrukturer.
De är karakteristiska helikaser av virus och har en unik 3'-5'-translokationspolaritet. De har endast fyra mycket konserverade sekvensmotiv och bildar ringstrukturer eller ringar.
De beskrevs först i bakterier och bakteriofager. De är en grupp replikerande eller replikerande helikaser.
De har en unik 5'-3'-translokationspolaritet och har fem mycket bevarade sekvensmotiv. Dessa helikaser kännetecknas av att forma ringar.
De är proteiner av typen Rho-faktor. Helikaserna i SF5-superfamiljen är karakteristiska för prokaryota organismer och är hexameriska ATP-beroende. De anses vara nära besläktade med SF4; dessutom har de ringformade och icke-ringformiga former.
De är proteiner som tydligen är relaterade till SF3-superfamiljen; SF6 presenterar emellertid en domän av ATPas-proteiner associerade med olika cellulära aktiviteter (AAA-proteiner) som inte finns i SF3.
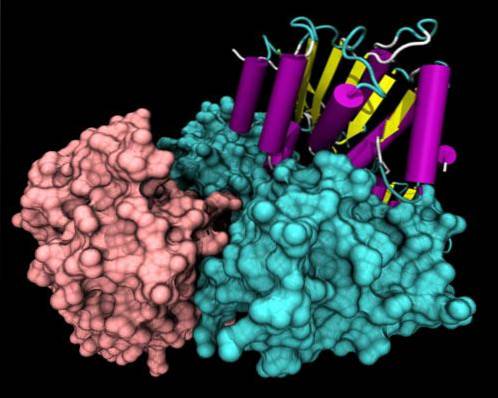
Strukturellt har alla helikaser mycket konserverade sekvensmotiv i den främre delen av sin primära struktur. En del av molekylen har ett särskilt aminosyrarrangemang som beror på den specifika funktionen hos varje helikas..
De mest strukturellt studerade helikaserna är de från SF1-superfamiljen. Det är känt att dessa proteiner grupperas i två domäner som liknar de multifunktionella RecA-proteinerna, och dessa domäner bildar en ATP-bindningsficka mellan dem..
Icke-konserverade regioner kan presentera specifika domäner av DNA-igenkänningstyp, cell-lokaliseringsdomän och protein-protein..
Funktionerna för dessa proteiner beror på en viktig mängd olika faktorer, inklusive miljöstress, celllinje, genetisk bakgrund och cellcykelstadier..
SF1 DNA-helikaserna är kända för att uppfylla specifika funktioner inom DNA-reparation, replikering, överföring och rekombination..
De separerar delar av en DNA-dubbelspiral och deltar i telomerunderhåll, dubbelsträngsreparation och avlägsnande av nukleinsyraassocierade proteiner.
Som tidigare nämnts är RNA-helikaser viktiga i de allra flesta av metaboliska RNA-processer, och det är också känt att dessa proteiner deltar i detektionen av viralt RNA..
Dessutom verkar de i det antivirala immunsvaret, eftersom de upptäcker främmande RNA eller främmande för kroppen (hos ryggradsdjur).
Hellicaser hjälper celler att övervinna endogen och exogen stress, undvika kromosomal instabilitet och upprätthålla cellulär balans.
Fel i detta system eller homeostatisk jämvikt är relaterat till genetiska mutationer som involverar gener som kodar proteiner av helikastyp; av denna anledning är de föremål för biomedicinska och genetiska studier.
Nedan kommer vi att nämna några av de sjukdomar som är relaterade till mutationer i gener som kodar DNA som helikastyp-proteiner:
Det är en genetisk sjukdom som orsakas av en mutation i en gen som heter WRN, som kodar för ett helikas. Mutant helikaset fungerar inte korrekt och orsakar ett antal sjukdomar som tillsammans utgör Werner syndrom.
Huvudegenskapen för dem som lider av denna patologi är deras för tidiga åldrande. För att sjukdomen ska manifestera sig måste den mutanta genen ärvas från båda föräldrarna. dess förekomst är mycket låg och det finns ingen behandling för dess botemedel.
Bloom syndrom är en genetisk sjukdom som härrör från mutationen av en autosomal gen som heter BLM som kodar för ett helikasprotein. Det förekommer bara för individer som är homozygota för den karaktären (recessiv).
Huvudegenskapen för denna sällsynta sjukdom är överkänslighet mot solljus, vilket orsakar hudskador på erytromatöst utslag. Det finns inget botemedel ännu.
Det är också känt som medfödd atrofisk poikiloderma. Det är en patologi av mycket sällsynt genetiskt ursprung: hittills finns det mindre än 300 fall som beskrivs över hela världen.
Det orsakas av en mutation i RECQ4-genen, en autosomal och recessiv gen som ligger på kromosom 8.
Symtom eller tillstånd hos detta syndrom inkluderar unga grå starr, abnormiteter i skelettsystemet, depigmentering, kapillärutvidgningar och atrofi i huden (poikiloderma). I vissa fall kan hypertyreoidism och brist på testosteronproduktion uppstå..


Ingen har kommenterat den här artikeln än.