A homozygot i genetik är det en individ som har två kopior av samma allel (samma version av en gen) i en eller flera loci (placera på kromosomen). Termen används ibland för större genetiska enheter såsom fullständiga kromosomer; i detta sammanhang är en homozygot en individ med två identiska kopior av samma kromosom.
Ordet homozygot består etymologiskt av två element. Villkoren är homo -lika eller identisk- och zygot -befruktat ägg eller den första cellen hos en individ uppstod genom sexuell reproduktion-.
Artikelindex
Organismer klassificeras baserat på olika egenskaper associerade med det genetiska materialet (DNA) som finns i deras celler. Med tanke på den cellulära strukturen där det genetiska materialet finns, har organismer klassificerats i två huvudtyper: prokaryoter (proffs: innan; Karyon: kärna) och eukaryoter (eu: Sann; Karyon: kärna).
I prokaryota organismer är det genetiska materialet begränsat till en viss region i cytoplasman hos celler som kallas nukleoid. Modellorganismerna i denna grupp motsvarar bakterier av arten Escherichia coli, som har en enda cirkulär DNA-sträng, det vill säga deras ändar är sammanfogade.
Denna kedja är känd som en kromosom och i E coli den innehåller cirka 1,3 miljoner baspar. Det finns några undantag från detta mönster inom gruppen, till exempel har vissa bakteriegener raka kedjekromosomer såsom spiroketer av släktet Borrelia.
Den linjära storleken eller längden av bakteriella genom / kromosomer ligger i allmänhet i intervallet millimeter, det vill säga de är flera gånger större än storleken på cellerna själva..
Genetiskt material lagras i en förpackad form för att minska utrymmet i denna stora molekyl. Denna packning uppnås genom supercoiling, en vridning på molekylens huvudaxel som ger små vändningar som, när de roterar, orsakar.
I sin tur, större vridningar av dessa små trådar på sig själva och resten av kedjan, vilket minskar avståndet och utrymmet mellan olika sektioner av den cirkulära kromosomen och leder den till en kondenserad form (vikt).
I eukaryoter är det genetiska materialet beläget i ett specialavdelning omgivet av ett membran; Detta fack är känt som cellens kärna..
Det genetiska materialet som finns i kärnan är strukturerat under en princip som liknar prokaryoter, super-lindningen.
Emellertid är graden / nivåerna av kinking högre eftersom mängden DNA att rymma är mycket högre. I eukaryoter innehåller kärnan inte en enda DNA-kedja eller kromosom, den innehåller flera av dem och dessa är inte cirkulära, utan linjära och måste ordnas.
Varje kromosom varierar i storlek beroende på art, men de är vanligtvis större än prokaryoter jämfört med varandra..
Till exempel är mänsklig kromosom 1 7,3 centimeter lång, medan kromosom av E coli mäter cirka 1,6 millimeter. För ytterligare referens innehåller det humana genomet 6,6 × 109 nukleotider.
Det finns en annan klassificering av organismer baserat på mängden genetiskt material de innehåller, känd som ploidi..
Organismer med en enda uppsättning eller kopia av kromosomer är kända som haploida (bakterier eller reproduktiva celler hos människor), med två uppsättningar / kopior av kromosomer är kända som diploida (Homo sapiens, Mus musculus, bland många andra), med fyra uppsättningar / kopior av kromosomer kallas de tetraploider (Odontophrinus americanus, växter av släktet Brassicca).
Organismer med ett stort antal kromosomsatser kallas kollektivt polyploider. I många fall är de extra kromosomerna kopior av en basuppsättning.
Under flera år ansågs att egenskaper som ploidi större än en var typiska för organismer med en definierad cellkärna, men de senaste resultaten har visat att vissa prokaryoter har flera kromosomala kopior som höjer sin ploidi, vilket framgår av fallen med Deinococcus radiodurans Y Bacillus meagateriium.
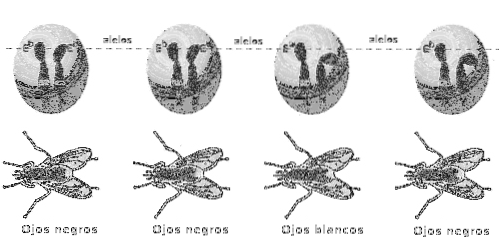
I diploida organismer (som ärtor studerade av Mendel) är de två generna av a loci, eller alleler, en ärvs genom moderen och den andra genom fadern och paret av alleler tillsammans representerar genotypen för den specifika genen.
En individ som presenterar en homozygot (homozygot) genotyp för en gen är en som har två identiska varianter eller alleler i en ställe tärningar.
Homozygos kan i sin tur delas in i två typer baserat på deras förhållande och bidrag till fenotypen: dominant och recessiv. Det bör noteras att båda uttrycken är fenotypiska egenskaper.
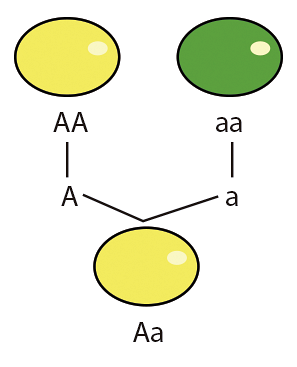
Dominans i genetiskt sammanhang är ett förhållande mellan allelerna i en gen där det fenotypiska bidraget från en allel maskeras av bidraget från den andra allelen av samma ställe; i detta fall är den första allelen recessiv och den andra är dominerande (heterozygositet).
Dominans ärvs inte i allelerna eller i fenotypen de producerar, det är ett förhållande som upprättas baserat på de närvarande allelerna och kan modifieras av externa medel som andra alleler..
Ett klassiskt exempel på dominans och dess förhållande till fenotypen är produktionen av ett funktionellt protein av den dominerande allelen som äntligen producerar den fysiska egenskapen, medan den recessiva allelen inte producerar proteinet i en funktionell form (mutant) och därför inte bidrar till fenotypen.
Således är en individuell homozygot dominant för en egenskap / egenskap en som har en genotyp som presenterar två identiska kopior av den dominerande allelen (ren linje).
Det är också möjligt att hitta dominans i genotyper där de två dominerande allelerna inte finns, men en dominerande allel är närvarande och en är recessiv, men detta är inte ett fall av homozygositet, det är ett fall av heterozygositet..
I genetisk analys representeras dominerande alleler av en stor bokstav relaterad till det drag som beskrivs..
I fallet med ärtblommans kronblad är den vilda egenskapen (i detta fall den lila färgen) dominerande och genotypen representeras som "P / P", vilket betecknar både den dominerande egenskapen och det homozygota tillståndet, det vill säga, närvaron av två identiska alleler i en diploid organism.
Å andra sidan bär en individuell homozygot recessiv för ett visst drag två kopior av allelen som kodar för den recessiva egenskapen.
Fortsätt med exemplet med ärten är den recessiva egenskapen i kronbladen den vita färgen, så hos individer med blommor av denna färg representeras varje allel med små bokstäver som antyder recessivitet och de två identiska recessiva kopiorna, så genotypen symboliseras som "p / p".
I vissa fall använder genetiker en stor bokstav symboliskt för att representera den vilda allelen (till exempel P) och därigenom symbolisera och hänvisa till en specifik nukleotidsekvens.
Å andra sidan, när en liten bokstav, p, används, representerar den en recessiv allel som kan vara vilken som helst av de möjliga typerna (mutationer) [1,4,9].
Processerna genom vilka en viss genotyp kan producera en fenotyp i organismer är varierad och komplex. Recessiva mutationer inaktiverar vanligtvis den drabbade genen och leder till funktionsförlust.
Detta kan hända genom ett partiellt eller fullständigt avlägsnande av genen, genom avbrott av genens uttryck eller genom förändring av strukturen för det kodade proteinet som äntligen förändrar dess funktion..
Å andra sidan producerar ofta dominerande mutationer en förstärkning av funktionen, kan öka aktiviteten för en given genprodukt eller ge en ny aktivitet till produkten och kan därför också producera olämpligt spatio-temporalt uttryck..
Denna typ av mutationer kan också associeras med funktionsförlust, det finns vissa fall där två kopior av en gen krävs för normal funktion så att avlägsnandet av en enda kopia kan leda till en mutant fenotyp..
Dessa gener är kända som haplo-otillräckliga. I vissa andra fall kan mutationen leda till strukturella förändringar i proteiner som stör funktionen hos vildtypsproteinet som kodas av den andra allelen. Dessa är kända som dominerande negativa mutationer. .
Hos människor är exempel på kända recessiva fenotyper albinism, cystisk fibros och fenylketonuri. Dessa är alla medicinska tillstånd med liknande genetiska baser..
Om vi tar det senare som ett exempel har individer med denna sjukdom en "p / p" -genotyp, och eftersom individen har båda recessiva alleler är den homozygot.
I detta fall är "p" relaterat till termen på engelska fenylketonuri y är gemener för att representera allelens recessiva karaktär. Sjukdomen orsakas av onormal bearbetning av fenylalanin, som under normala förhållanden bör omvandlas till tyrosin (båda molekylerna är aminosyror) av enzymet fenylalaninhydroxylas..
Mutationer i närheten av det aktiva stället för detta enzym hindrar det från att kunna binda till fenylalanin för att senare bearbeta det.
Som en följd ackumuleras fenylalanin i kroppen och omvandlas till fenylpyruvsyra, en förening som stör nervsystemets utveckling. Dessa tillstånd är kollektivt kända som autosomala recessiva störningar..
Arvsmönstren och därför närvaron av alleler för en gen, både dominerande och recessiv, i genotyperna av individer inom en befolkning följer Mendels första lag.
Denna lag är känd som lagen om jämlik segregering av alleler och har molekylära baser som förklaras under bildandet av könsceller..
I diploida organismer som reproducerar sexuellt finns det två huvudcelltyper: somatiska celler och könsceller eller könsceller..
Somatiska celler har två kopior av varje kromosom (diploid) och var och en av kromosomerna (kromatiderna) innehåller en av de två allelerna.
Gametiska celler produceras av groddvävnader genom meios där diploida celler genomgår kärnuppdelning åtföljd av en kromosomreduktion under denna process, följaktligen har de bara en uppsättning kromosomer, så de är haploida.
Under meios är den akromatiska spindeln förankrad i kromosomernas centromerer och kromatiderna separeras (och därför allelerna också) mot motsatta poler i stamcellen, vilket ger två separata dotterceller eller gameter..
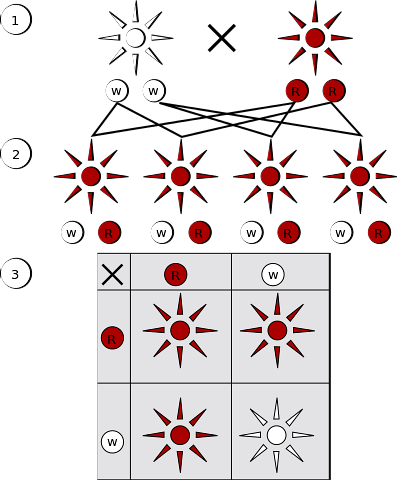
Om individen som producerar könscellerna är homozygot (A / A eller a / a) kommer det totala antalet gametiska celler som produceras av honom att ha identiska alleler (A respektive a).
Om individen är heterozygot (A / a eller a / A) kommer hälften av könscellerna att ha en allel (A) och den andra halvan den andra (a). När sexuell reproduktion är klar bildas en ny zygot, de manliga och kvinnliga könscellerna smälter samman för att bilda en ny diploid cell och ett nytt par kromosomer och därför upprättas alleler..
Denna process har sitt ursprung i en ny genotyp som bestäms av allelerna som bidragits av den manliga könscellen och den kvinnliga könscellen..
I mendelisk genetik har homozygota och heterozygota fenotyper inte samma sannolikhet att de kommer att visas i en population, men de möjliga allelkombinationer som är associerade med fenotyperna kan härledas eller bestämmas genom genetisk korsanalys..
Om båda föräldrarna är homozygota för en gen av den dominerande typen (A / A), kommer båda könscellerna att vara av typ A i sin helhet och deras förening kommer alltid att resultera i en A / A-genotyp..
Om båda föräldrarna har en homozygot recessiv genotyp (a / a), kommer avkomman alltid att resultera i en homozygot recessiv genotyp också..
I evolutionsteorin sägs det att utvecklingsmotorn är förändring och på genetisk nivå sker förändring genom mutationer och rekombinationer.
Mutationer involverar ofta förändringar i någon nukleotidbas av en gen, även om de kan ha mer än en bas.
De flesta mutationer betraktas som spontana händelser associerade med felfrekvensen eller trohet hos polymeraser under DNA-transkription och replikering..
Det finns också mycket bevis på fysiska fenomen som orsakar mutationer på genetisk nivå. För sin del kan rekombinationer producera utbyte av hela sektioner av kromosomer men är bara associerade med händelser av celldubblering, såsom mitos och meios.
Faktum är att de anses vara en grundläggande mekanism för att generera genotypisk variation under gametbildning. Införlivande av genetisk variation är ett kännetecken för sexuell reproduktion.
Fokuserad på gener anses det för närvarande att arvsmotorn och därmed evolutionen är gener som presenterar mer än en allel.
De gener som bara har en allel kan knappast orsaka en evolutionär förändring om alla individer i befolkningen har två kopior av samma allel som exemplifierats ovan..
Detta beror på att när genetisk information överförs från en generation till en annan kommer förändringar knappast att hittas i den populationen om det inte finns krafter som producerar variationer i gener som de som nämns ovan också..
De enklaste evolutionära modellerna är de som bara överväger a ställe och dess mål är att försöka förutsäga genotypfrekvenserna i nästa generation, från data från den befintliga generationen.


Ingen har kommenterat den här artikeln än.