
De mitotisk spindel eller akromatisk, även kallad mitotisk maskineri, är en cellulär struktur som består av mikrotubuli av proteinnatur som bildas under celldelning (mitos och meios).
Uttrycket akromatisk hänvisar till det faktum att det inte fläckar med färgämnena Ace- eller B. Spindeln deltar i en rättvis fördelning av genetiskt material mellan de två dottercellerna, som härrör från celldelning.
Celldelning är den process genom vilken både könscellerna, som är meiotiska celler, och de somatiska cellerna som är nödvändiga för tillväxt och utveckling av en organism genereras från zygoten..
Övergången mellan två på varandra följande uppdelningar utgör cellcykeln, vars varaktighet varierar mycket beroende på celltyp och stimuli som den utsätts för..
Under mitosen av en eukaryot cell (cell som har en äkta kärna och organeller avgränsade av membran) uppstår flera faser: S-fas, profas, prometafas, metafas, anafas, telofas och gränssnitt.
Ursprungligen kondenserar kromosomerna och bildar två identiska filament som kallas kromatider. Varje kromatid innehåller en av de två tidigare genererade DNA-molekylerna, sammanfogade av en region som kallas centromeren, som spelar en grundläggande roll i migrationsprocessen mot polerna före celldelning..
Mitotisk uppdelning sker under en organisms liv. Det uppskattas att cirka 10 inträffar i kroppen under människolivet17 celldelningar. Meiotisk uppdelning sker i könsceller som producerar könet eller könsceller.
Artikelindex
Den akromatiska spindeln anses vara ett längsgående system av proteinmikrofibriller eller cellulära mikrotubuli. Den bildas vid celldelningen mellan de kromosomala centromererna och centrosomerna vid cellpolerna och är relaterad till migrationen av kromosomer för att generera dotterceller med samma mängd genetisk information.
Centrosomen är regionen där mikrotubuli kommer från både den akromatiska spindeln och cytoskelettet. Dessa spindelmikrotubuli består av tubulin-dimerer som lånas från cytoskelettet..
I början av mitosen avviker mikrotubuli-nätverket i cellens cytoskelett och den akromatiska spindeln bildas. Efter att celldelning inträffar, artikuleras spindeln och mikrotubuli-nätverket i cytoskelettet omorganiseras och återställer cellen till sitt vilotillstånd.
Det är viktigt att skilja på att det finns tre typer av mikrotubuli i den mitotiska apparaten: två typer av spindelmikrotubuli (kinetochore och polära mikrotubuli) och en typ av aster mikrotubuli (astrala mikrotubuli).
Den bilaterala symmetrin för den akromatiska spindeln beror på interaktioner som håller sina två halvor samman. Dessa interaktioner är: antingen laterala, mellan de överlappande positiva ändarna av de polära mikrotubuli; eller de är terminala interaktioner mellan mikrotubuli i kinetochore och kinetochore hos systerkromatiderna.
DNA-replikering inträffar under S-fasen i cellcykeln, sedan, under profas, uppträder migrationen av centrosomerna till motsatta poler i cellen och kromosomerna kondenserar också.
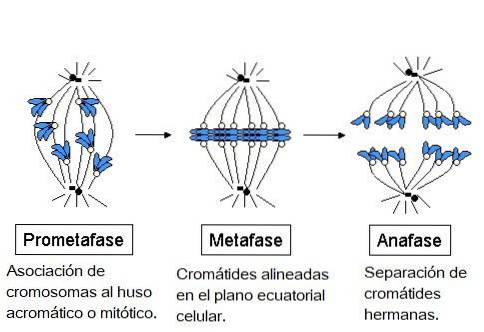
I prometafasen sker bildandet av det mitotiska maskineriet tack vare sammansättningen av mikrotubuli och deras penetration i det inre av kärnan. Systerkromatider länkade av centromererna genereras och dessa i sin tur binder till mikrotubuli.
Under metafas inriktas kromosomerna i cellens ekvatoriella plan. Spindeln är organiserad i en central mitotisk spindel och ett par asters.
Varje aster består av mikrotubuli anordnade i en stjärnform som sträcker sig från centrosomerna till cellbarken. Dessa astrala mikrotubuli interagerar inte med kromosomer.
Det sägs sedan att aster strålar från centrosomen till cellbarken och deltar både i placeringen av hela mitotiska apparaten och vid bestämning av planet för celldelning under cytokinese..
Senare, under anafas, är mikrotubuli i den akromatiska spindeln förankrad av en positiv ände på kromosomerna genom sina kinetochores och av en negativ ände till en centrosom.
Separation av systerkromatider i oberoende kromosomer sker. Varje kromosom som är fäst vid en kinetochore mikrotubuli rör sig mot en cellpol. Samtidigt sker separationen av cellpolerna.
Slutligen, under telofas och cytokinese, bildas kärnmembran runt dotterkärnor och kromosomer förlorar sitt kondenserade utseende..
Den mitotiska spindeln försvinner när mikrotubuli avpolymeriseras och celldelning sker in i gränssnittet.
Mekanismen som är involverad i migrationen av kromosomerna mot polerna och den efterföljande separationen av polerna från varandra är dock inte exakt känd; Det är känt att interaktioner mellan kinetochore och mikrotubuli i spindeln fäst vid den är involverade i denna process..
När varje kromosom migrerar mot motsvarande pol inträffar depolymerisation av den bifogade mikrotubuli, eller kinetokorisk mikrotubuli. Man tror att denna depolymerisation kan generera den passiva rörelsen för kromosomen fäst vid mikrotubuli i spindeln..
Man tror också att det kan finnas andra motorproteiner associerade med kinetokoren, i vilken energin från hydrolysen av ATP skulle användas..
Denna energi skulle tjäna till att driva kromosomens migrering längs mikrotubuli till dess ände som kallas "mindre" där centrosomen är belägen..
Samstämmigt kan depolymerisationen av änden av mikrotubuli som ansluter sig till kinetokoren eller "plus" -änden inträffa, vilket också skulle bidra till kromosomens rörelse.
Den akromatiska eller mitotiska spindeln är en cellulär struktur som uppfyller funktionen att förankra kromosomerna genom deras kinetokorer, rikta in dem med cellekvatorn och slutligen styra migrationen av kromatiderna mot de motsatta polerna i cellen före deras uppdelning, vilket möjliggör fördelningen genetiskt material mellan de två resulterande dottercellerna.
Om fel uppstår i denna process genereras brist eller överskott av kromosomer, vilket översätts till onormala utvecklingsmönster (inträffar under embryogenes) och olika patologier (inträffar efter individens födelse).
Det finns bevis för att mikrotubuli i den akromatiska spindeln deltar i att bestämma placeringen av de strukturer som är ansvariga för cytoplasmatisk uppdelning..
Det viktigaste beviset är att celldelning alltid sker i spindelns mittlinje, där polära fibrer överlappar varandra..
Evolutionärt har det valts ut som en mycket överflödig mekanism, i vilken varje steg utförs av mikrotubuli-motorproteiner.
Man tror att det evolutionära förvärvet av mikrotubuli berodde på en process av endosymbios, i vilken en eukaryot cell absorberade en prokaryot cell som uppvisade dessa akromatiska spindelstrukturer från omgivningen. Allt detta kunde ha hänt innan mitosen började.
Denna hypotes antyder att mikrotubuli-proteinstrukturerna ursprungligen kunde ha uppfyllt en framdrivningsfunktion. Då mikrotubuli blev en del av en ny organism, skulle de utgöra cytoskelettet och senare det mitotiska maskineriet.
I evolutionär historia har det förekommit variationer i grundschemat för eukaryot celldelning. Celldelning representerade endast några faser av cellcykeln, vilket är en stor process.



Ingen har kommenterat den här artikeln än.