De mitos det är en process med celldelning, där en cell producerar genetiskt identiska dotterceller; för varje cell genereras två "döttrar" med samma kromosombelastning. Denna uppdelning äger rum i de somatiska cellerna i eukaryota organismer..
Denna process är ett av stegen i cellcykeln för eukaryota organismer, som består av fyra faser: S (DNA-syntes), M (celldelning), G1 och G2 (mellanfaser där mRNA och proteiner produceras). Tillsammans betraktas faserna G1, G2 och S som ett gränssnitt. Kärn- och cytoplasmatisk uppdelning (mitos och cytokinese) utgör det sista steget i cellcykeln.
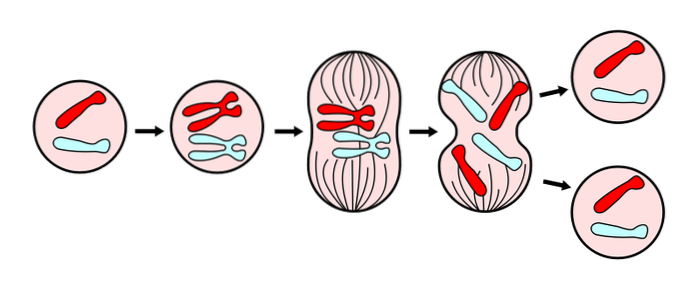
På molekylär nivå initieras mitos genom aktivering av ett kinas (protein) som kallas MPF (Maturation Promoting Factor) och den därav följande fosforyleringen av ett betydande antal av cellens komponentproteiner. Det senare gör att cellen kan presentera de morfologiska förändringar som är nödvändiga för att genomföra delningsprocessen..
Mitos är en asexuell process, eftersom stamcellen och dess döttrar har exakt samma genetiska information. Dessa celler är kända som diploida eftersom de bär hela kromosombelastningen (2n).
Meios är å andra sidan processen för celldelning som leder till sexuell reproduktion. I denna process replikerar en diploid stamcell sina kromosomer och delar sig sedan två gånger i rad (utan att replikera dess genetiska information). Slutligen genereras 4 dotterceller med bara hälften av kromosombelastningen, som kallas haploida (n).
Artikelindex
Mitos i encelliga organismer producerar vanligtvis dotterceller som mycket liknar deras förfäder. Däremot kan denna process under utvecklingen av flercelliga varelser ge upphov till två celler med vissa olika egenskaper (trots att de är genetiskt identiska)..
Denna celldifferentiering ger upphov till de olika celltyper som utgör flercelliga organismer..
Under en organisms liv sker cellcykeln kontinuerligt och bildar ständigt nya celler som i sin tur växer och förbereder sig för att dela sig genom mitos.
Celltillväxt och delning regleras av mekanismer, såsom apoptos (programmerad celldöd), som gör det möjligt att upprätthålla en balans, vilket undviker överflöd av vävnad. På detta sätt säkerställs att de defekta cellerna ersätts av nya celler, i enlighet med kroppens krav och behov..
Förmågan att reproducera är en av de viktigaste egenskaperna hos alla organismer (från unicellular till multicellular) och av cellerna som komponerar den. Denna kvalitet säkerställer kontinuiteten i din genetiska information..
Att förstå processerna för mitos och meios har spelat en grundläggande roll för att förstå organismernas spännande cellulära egenskaper. Till exempel egenskapen att hålla antalet kromosomer konstant från en cell till en annan inom en individ och mellan individer av samma art.
När vi lider av någon form av skär eller sår på vår hud, observerar vi hur den skadade huden på några dagar återhämtar sig. Detta inträffar tack vare processen med mitos.
I allmänhet följer mitos samma sekvens av processer (faser) i alla eukaryota celler. I dessa faser inträffar många morfologiska förändringar i cellen. Bland dem kondensering av kromosomer, bristning av kärnmembranet, separation av cellen från den extracellulära matrisen och andra celler och uppdelningen av cytoplasman..
I vissa fall anses kärnkraftsdelning och cytoplasmatisk uppdelning vara distinkta faser (mitos respektive cytokinese).
För en bättre studie och förståelse av processen har sex (6) faser utsetts, kallade: profas, prometafas, metafas, anafas och telofas, med beaktande av cytokinese som en sjätte fas, som börjar utvecklas under anafas..
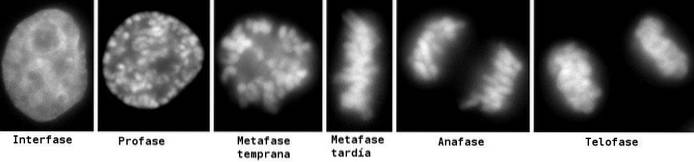
Dessa faser har studerats sedan 1800-talet genom ljusmikroskopet, så idag är de lätt igenkännliga enligt de morfologiska egenskaper som cellen presenterar, såsom kromosomkondensation och bildandet av den mitotiska spindeln..
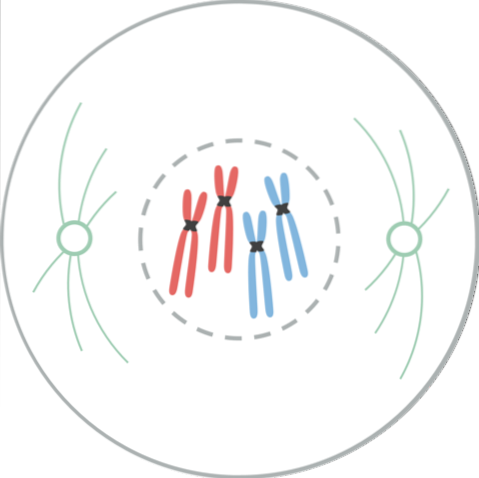
Profas är den första synliga manifestationen av celldelning. I denna fas kan utseendet på kromosomer ses som urskiljbara former på grund av den progressiva komprimeringen av kromatin. Denna kondensation av kromosomer börjar med fosforylering av Histone H1-molekyler med MPF-kinas..
Kondensprocessen består av sammandragningen och därför minskningen av kromosomernas storlek. Detta inträffar på grund av kromatinfibrernas lindning, vilket ger lättare förskjutbara strukturer (mitotiska kromosomer).
Kromosomer som tidigare duplicerats under cellcykelns S-period, får ett dubbelsträngat utseende, så kallade systerkromatider, dessa strängar hålls samman genom en region som kallas centromeren. I denna fas försvinner också nukleolierna.
Under profas bildas den mitotiska spindeln, som består av mikrotubuli och proteiner som utgör en uppsättning fibrer.
När spindeln bildas demonteras mikrotubuli i cytoskelettet (genom att avaktivera proteinerna som bibehåller deras struktur), vilket ger det nödvändiga materialet för bildandet av nämnda mitotiska spindel..
Centrosomen (en membranfri organell, funktionell i cellcykeln), duplicerad vid gränssnittet, fungerar som monteringsenheten för spindelmikrotubuli. I djurceller har centrosomen ett par centrioler i mitten; men dessa saknas i de flesta växtceller.
De duplicerade centrosomerna börjar separera från varandra medan mikrotubuli i spindeln är monterade i var och en av dem och börjar migrera mot motsatta ändar av cellen..
I slutet av profaset börjar kärnhuvudets bristning, som inträffar i separata processer: demontering av kärnporer, nukleära lameller och kärnmembran. Denna brytning gör att den mitotiska spindeln och kromosomerna kan börja interagera..
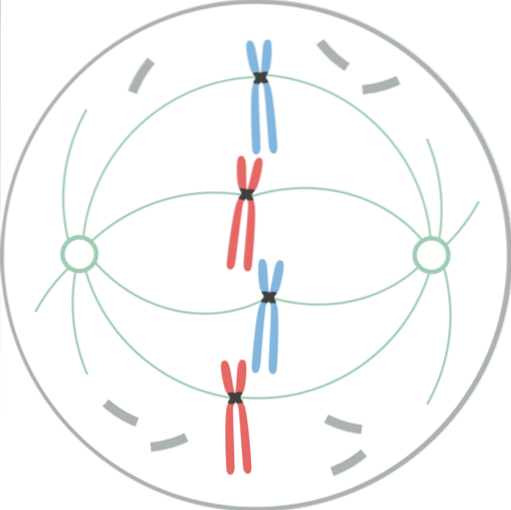
I detta skede har kärnhuvudet varit helt fragmenterat, så spindelns mikrotubuli invaderar detta område och interagerar med kromosomerna. De två centrosomerna har separerats, vardera belägna vid polerna i den mitotiska spindeln, i motsatta ändar av cellerna..
Nu innefattar den mitotiska spindeln mikrotubuli (som sträcker sig från varje centrosom mot mitten av cellen), centrosomerna och ett par asters (strukturer med en radiell fördelning av korta mikrotubuli, som utvecklas från varje centrosom)..
Kromatiderna utvecklade var och en en specialiserad proteinstruktur, kallad kinetochore, som ligger i centromeren. Dessa kinetokorer är placerade i motsatta riktningar och vissa mikrotubuli, kallade mikrotubuli i kinetochore, följer dem..
Dessa mikrotubuli, fästa vid kinetokoren, börjar röra sig till kromosomen från den ände de sträcker sig; några från en pol och andra från motsatt pol. Detta skapar en "dra och krympa" -effekt som, när den är stabiliserad, gör att kromosomen hamnar mellan celländarna..
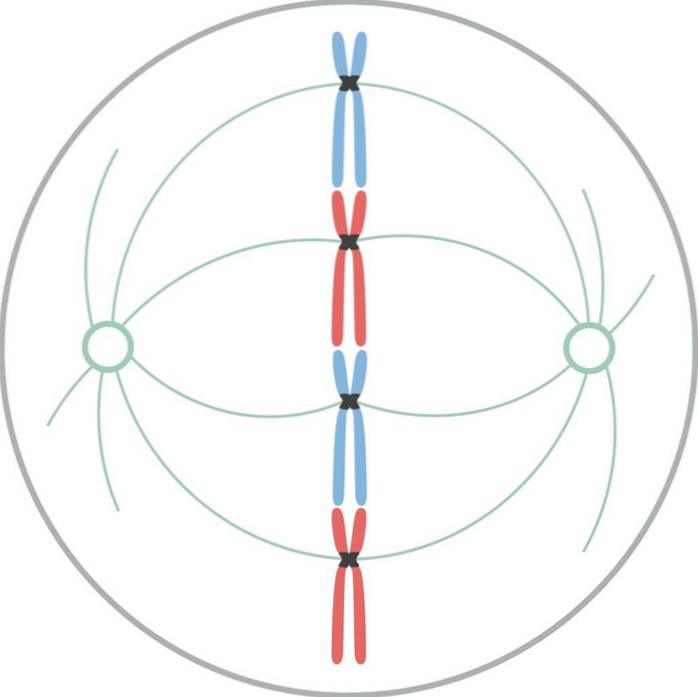
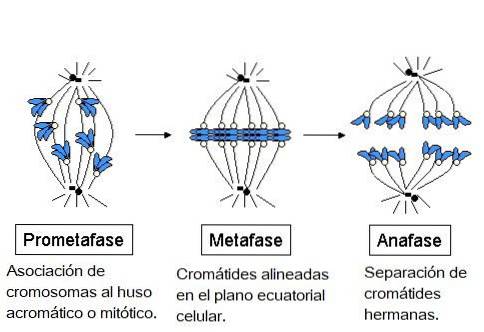
I metafas är centrosomer placerade i motsatta ändar av celler. Spindeln visar en tydlig struktur, i vilken kromosomerna ligger. Centromererna i dessa kromosomer är fästa vid fibrerna och inriktade i ett imaginärt plan som kallas metafasplattan..
Kromatidkinetochoresna förblir fästa vid mikrotubuli i kinetochore. Mikrotubuli som inte fäster vid kinetokorerna och sträcker sig från motsatta poler av spindeln interagerar nu med varandra. Vid denna punkt är mikrotubuli från asterna i kontakt med plasmamembranet..
Denna tillväxt och interaktion mellan mikrotubuli kompletterar den mitotiska spindelns struktur och ger den ett "fågelbur" -utseende..
Morfologiskt är denna fas den med minst förändringar, varför den ansågs vara en vilopas. Men även om de inte är lätt märkbara, förekommer många viktiga processer i det, förutom att det är det längsta stadiet av mitos.
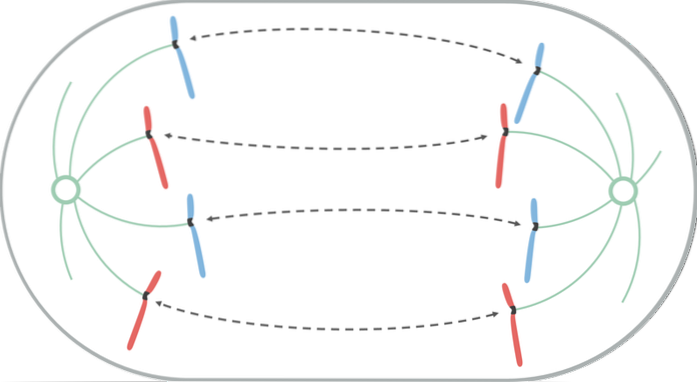
Under anafas börjar varje kromatidpar separera (på grund av inaktivering av proteinerna som håller dem ihop). Separerade kromosomer rör sig till motsatta ändar av cellen.
Denna migrationsrörelse beror på förkortningen av kinetokorns mikrotubuli, vilket genererar en "drag" -effekt som får varje kromosom att röra sig från sin centromer. Beroende på placeringen av centromeren på kromosomen kan den ta en viss V- eller J-form under dess förskjutning..
Mikrotubuli som inte fästs vid kinetokoren, växer och förlänger sig genom vidhäftningen av tubulin (protein) och genom verkan av motorproteiner som rör sig över dem, vilket gör att kontakten mellan dem kan stoppas. När de rör sig bort från varandra gör spindelns poler också och förlänger cellen.
I slutet av denna fas är kromosomgrupperna placerade i motsatta ändar av den mitotiska spindeln och lämnar varje ände av cellen med en komplett och ekvivalent uppsättning kromosomer..
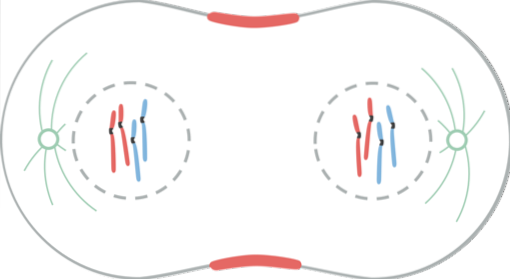
Telofasen är den sista fasen av kärnkraftsuppdelningen. Kinetochore-mikrotubuli sönderfaller medan polära mikrotubuli förlängs ytterligare.
Kärnmembranet börjar bildas runt varje kromosomuppsättning med hjälp av kärnhöljen hos stamcellen, som var som vesiklar i cytoplasman.
I detta steg dekondenseras kromosomerna som är vid cellpolerna helt på grund av defosforylering av histonmolekylerna (H1). Bildningen av kärnmembranets element styrs av flera mekanismer.
Under anafas började många av de fosforylerade proteinerna i profas att avfosforylera. Detta gör att i början av telofasen börjar kärnblåsorna återmonteras och associeras med kromosomens yta..
Å andra sidan är kärnporerna återmonterade så att kärnproteiner kan pumpas. Proteinerna i kärnplattan avfosforyleras, så att de kan associeras igen, för att slutföra bildningen av kärnplattan.
Slutligen, efter att kromosomerna är helt dekondenserade, startas RNA-syntesen igen, bildar nukleolierna igen och därmed fullbordas bildningen av de nya interfaskärnorna i dottercellerna..

Cytokinese tas som en separat händelse från kärnkraftsdelning, och i typiska celler följer den cytoplasmiska delningsprocessen varje mitos, med början vid anafas. Flera studier har visat att i vissa embryon uppstår flera nukleära uppdelningar före cytoplasmatisk uppdelning.
Processen börjar med uppkomsten av ett spår eller en klyft som är markerad i metafasplattans plan, vilket säkerställer att uppdelningen sker mellan kromosomgrupperna. Platsen för klyftan indikeras av den mitotiska spindeln specifikt, asternas mikrotubuli.
I den markerade klyftan finns en serie mikrofilament som bildar en ring riktad mot den cytoplasmiska sidan av cellmembranet, till stor del sammansatt av aktin och myosin. Dessa proteiner interagerar med varandra så att ringen kan dra ihop sig runt spåret..
Denna sammandragning alstras genom glidningen av filamenten hos dessa proteiner, när de interagerar med varandra, på samma sätt som de gör, till exempel i muskelvävnader..
Kontraktionen av ringröret fördjupas och utövar en "klämeffekt" som slutligen delar upp stamcellen, vilket möjliggör separering av dottercellerna med deras utvecklande cytoplasmiska innehåll..
Växtceller har en cellvägg, så deras cytoplasmiska uppdelningsprocess skiljer sig från den tidigare beskrivna och börjar i telofas..
Bildandet av en ny cellvägg börjar när mikrotubuli i den återstående spindeln monteras och utgör fragmoplasten. Denna cylindriska struktur består av två uppsättningar mikrotubuli som är anslutna i sina ändar och vars positiva poler är inbäddade i ett elektroniskt kort i ekvatorialplanet..
Små vesiklar från Golgi-apparaten, packade med cellväggsprekursorer, färdas genom fragmoplastens mikrotubuli till ekvatorialområdet och kombineras för att bilda en cellplatta. Innehållet i blåsorna utsöndras i denna platta när den växer.
Denna plack växer och smälter samman med plasmamembranet längs cellomkretsen. Detta sker på grund av den konstanta omläggningen av fragmoplastens mikrotubuli vid periferin av plattan, vilket gör att fler blåsor kan röra sig mot detta plan och tömma deras innehåll..
På detta sätt sker den cytoplasmiska separationen av dottercellerna. Slutligen tillåter innehållet i cellplattan tillsammans med cellulosamikrofibrerna i den att fullborda bildandet av den nya cellväggen..
Mitos är en mekanism för delning i celler och är en del av en av faserna i cellcykeln i eukaryoter. På ett enkelt sätt kan vi säga att huvudfunktionen för denna process är reproduktionen av en cell i två dotterceller.
För encelliga organismer betyder celldelning generering av nya individer, medan för multicellulära organismer är denna process en del av tillväxten och korrekt funktion av hela organismen (celldelning genererar vävnadsutveckling och underhåll av strukturer).
Mitosprocessen aktiveras enligt organismens krav. I däggdjur börjar till exempel röda blodkroppar (erytrocyter) dela sig och bildar fler celler när kroppen behöver bättre syreupptag. På samma sätt reproducerar vita blodkroppar (leukocyter) när det är nödvändigt att bekämpa en infektion..
Däremot saknar vissa specialiserade djurceller praktiskt taget mitosprocessen eller är det mycket långsamt. Exempel på detta är nervceller och muskelceller).
I allmänhet är de celler som ingår i kroppens bindväv och strukturvävnad och vars reproduktion endast är nödvändig när en cell har en defekt eller försämring och behöver bytas ut.
Systemet för celldelning och tillväxt är mycket mer komplext i flercelliga organismer än i encelliga. I det senare är reproduktion i princip begränsad av tillgången på resurser.
I djurceller arresteras delning tills det finns en positiv signal som aktiverar denna process. Denna aktivering kommer i form av kemiska signaler från närliggande celler. Detta gör det möjligt att förhindra obegränsad tillväxt av vävnader och reproduktion av defekta celler, vilket allvarligt kan skada organismens liv.
En av mekanismerna som styr cellmultiplikation är apoptos, där en cell dör (på grund av produktionen av vissa proteiner som aktiverar självförstörelse) om den uppvisar betydande skada eller är infekterad av ett virus.
Det finns också reglering av cellutveckling genom inhibering av tillväxtfaktorer (såsom proteiner). Således förblir cellerna vid gränssnittet utan att gå vidare till M-fasen i cellcykeln..
Mitosprocessen äger rum i de allra flesta eukaryota celler, från encelliga organismer som jäst, som använder den som en asexuell reproduktionsprocess, till komplexa flercelliga organismer som växter och djur..
Även om cellcykeln i allmänhet är densamma för alla eukaryota celler, finns det anmärkningsvärda skillnader mellan encelliga och flercelliga organismer. I det förra favoriseras tillväxt och delning av celler av naturligt urval. I flercelliga organismer är spridning begränsad av strikta kontrollmekanismer.
I encelliga organismer sker reproduktion på ett accelererat sätt, eftersom cellcykeln fungerar ständigt och dottercellerna snabbt går in mot mitos för att fortsätta med denna cykel. Medan cellerna i flercelliga organismer tar betydligt längre tid att växa och dela sig.
Det finns också vissa skillnader mellan de mitotiska processerna hos växt- och djurceller, som i vissa av faserna av denna process, men i princip fungerar mekanismen på liknande sätt i dessa organismer..
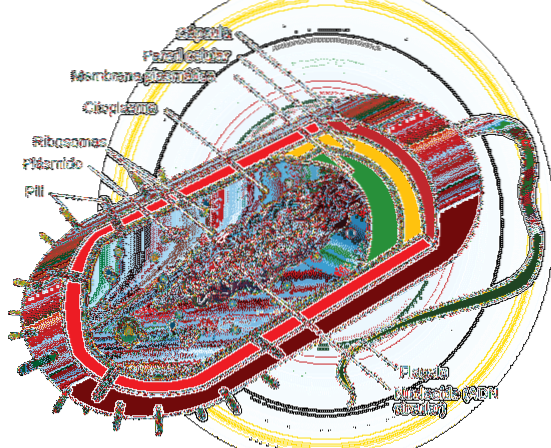
Prokaryota celler växer i allmänhet och delar sig snabbare än eukaryota celler.
Organismer med prokaryota celler (i allmänhet encelliga eller i vissa fall multicellulära) saknar ett kärnmembran som isolerar det genetiska materialet i en kärna, så det sprids i cellen, i ett område som kallas nukleoid. Dessa celler har en cirkulär huvudkromosom.
Celldelning i dessa organismer är således mycket mer direkt än i eukaryota celler, utan den beskrivna mekanismen (mitos). I dem utförs reproduktion med en process som kallas binär fission, där DNA-replikering börjar vid ett specifikt ställe på den cirkulära kromosomen (replikationsursprung eller OriC)..
Två ursprung bildas sedan som migrerar till motsatta sidor av cellen när replikering sker, och cellen sträcker sig till dubbelt så stor som den. I slutet av replikationen växer cellmembranet in i cytoplasman och delar stamcellen i två döttrar med samma genetiska material.
Utvecklingen av eukaryota celler förde med sig en ökning av genomets komplexitet. Detta involverade utvecklingen av mer detaljerade delningsmekanismer.
Det finns hypoteser som föreslår att bakteriedeling är föregångarmekanismen för mitos. Ett visst samband har hittats mellan proteiner associerade med binär fission (som kan vara de som förankrar kromosomer till specifika platser på plasmamembranet hos döttrar) med tubulin och aktin i eukaryota celler..
Vissa studier indikerar vissa särdrag i uppdelningen av moderna encelliga protister. I dem förblir kärnmembranet intakt under mitos. De replikerade kromosomerna förblir förankrade på vissa platser på detta membran och separerar när kärnan börjar sträcka sig under celldelning..
Detta visar viss sammanfall med processen för binär klyvning, där de replikerade kromosomerna fäster vid vissa platser på cellmembranet. Hypotesen föreslår sedan att protisterna som presenterar denna kvalitet under sin celldelning kunde ha bibehållit denna egenskap hos en förfäders prokaryot cell..
För närvarande har förklaringar ännu inte utvecklats till varför det i eukaryota celler av flercelliga organismer är nödvändigt att kärnmembranet sönderfaller under celldelningsprocessen..



Ingen har kommenterat den här artikeln än.