De kärnkärna det är en cellulär struktur som inte avgränsas av ett membran, och är en av de mest framträdande områdena i kärnan. Det ses som en tätare region i kärnan och är indelad i tre regioner: tät fibrillär komponent, fibrillär centrum och granulär komponent..
Det är huvudsakligen ansvarigt för syntes och sammansättning av ribosomer; denna struktur har dock också andra funktioner. Mer än 700 proteiner har hittats i kärnan som inte är involverade i ribosombiogenesprocesser. På samma sätt är kärnan involverad i utvecklingen av olika patologier.
Den första forskaren som observerade nukleolzonen var F. Fontana 1781, för mer än två århundraden sedan. Sedan i mitten av 1930-talet kunde McClintock observera en sådan struktur i sina experiment med Zea mays. Sedan dess har hundratals utredningar fokuserat på att förstå funktionerna och dynamiken i denna kärnregion..
Artikelindex
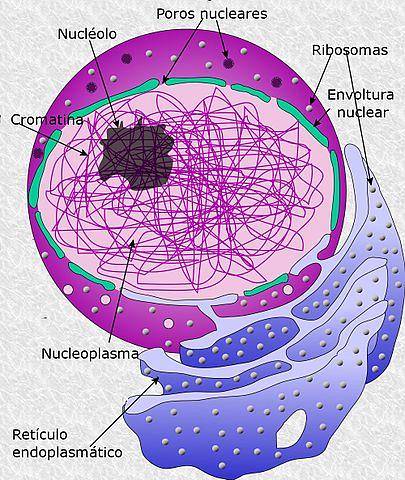
Kärnkärnan är en framträdande struktur belägen i kärnan i eukaryota celler. Det är en "region" i form av en sfär, eftersom det inte finns någon typ av biomembran som skiljer den från resten av kärnkomponenterna..
Det kan ses under mikroskopet som en delregion av kärnan när cellen är vid gränssnittet.
Det är organiserat i regioner som kallas NOR (för dess förkortning på engelska: kromosomala nukleolära organisatorregionerdär de ribosomkodande sekvenserna finns.
Dessa gener finns i specifika regioner i kromosomerna. Hos människor är de organiserade i tandem i satellitregionerna i kromosomerna 13, 14, 15, 21 och 22.
I kärnan sker transkription, bearbetning och montering av underenheterna som utgör ribosomerna.
Förutom sin traditionella funktion är kärnan relaterad till tumörundertryckande proteiner, cellcykelregulatorer och till och med proteiner från virus.
Nucleolus-proteiner är dynamiska och deras sekvens verkar ha bevarats under hela evolutionen. Av dessa proteiner har endast 30% associerats med ribosombiogenes.
Kärnan är uppdelad i tre huvudkomponenter, som kan särskiljas genom elektronmikroskopi: den täta fibrillära komponenten, den fibrillära mitten och den granulära komponenten..
I allmänhet är det omgivet av kondenserat kromatin, kallat heterokromatin. I kärnan sker processerna för transkription av ribosomalt RNA, bearbetning och sammansättning av ribosomala prekursorer.
Nukleolus är en dynamisk region, där proteinerna som komponenterna snabbt kan associera och separera från nukleolära komponenter, vilket skapar ett kontinuerligt utbyte med nukleoplasman (inre gelatinös substans i kärnan).
Hos däggdjur varierar strukturen av kärnan med stadierna i cellcykeln. I profas observeras en desorganisering av kärnan och den återmonteras i slutet av den mitotiska processen. Den maximala transkriptionsaktiviteten i kärnan har observerats i S- och G2-faserna.
Aktiviteten för RNA-polymeras I kan påverkas av olika tillstånd av fosforylering, vilket modifierar aktiviteten hos kärnan under cellcykeln. Tystnad under mitos inträffar på grund av fosforylering av olika element såsom SL1 och TTF-1.
Detta mönster är dock inte vanligt i alla organismer. Till exempel, i jäst är kärnan närvarande -och aktiv- under hela processen med celldelning..
Generna som kodar för ribosomalt RNA finns i fibrillära centra. Dessa centra är klara regioner omgivna av de täta fibrillära komponenterna. De fibrillära centren varierar i storlek och antal, beroende på celltyp.
Ett visst mönster har beskrivits med avseende på egenskaperna hos fibrillära centra. Celler med hög ribosomsyntes har ett lågt antal fibrillära centra, medan celler med reducerad metabolism (såsom lymfocyter) har större fibrillära centra..
Det finns specifika fall, såsom neuroner med en mycket aktiv metabolism, vars kärnkärna har ett jätte fibrillärt centrum, åtföljd av små mindre centra..
Den täta fibrillära komponenten och fibrillära centra är inbäddade i den granulära komponenten, vars granuler har en diameter av 15-20 nm. Transkriptionsprocessen (passage av DNA-molekylen till RNA, anses vara det första steget för genuttryck) sker vid gränserna för fibrillära centra och i den täta fibrillära komponenten.
Bearbetning av ribosomalt pre-RNA sker i den täta fibrillära komponenten och processen sträcker sig till den granulära komponenten. Avskrifter ackumuleras i den täta fibrillära komponenten och nukleolära proteiner finns också i den täta fibrillära komponenten. Det är i denna region där samlingen av ribosomer sker.
Efter att denna monteringsprocess av ribosomalt RNA med nödvändiga proteiner är avslutad exporteras dessa produkter till cytoplasman.
Den granulära komponenten är rik på transkriptionsfaktorer (SUMO-1 och Ubc9 är några exempel). Typiskt omges kärnan av heterokromatin; detta komprimerade DNA tros spela en roll i ribosomalt RNA-transkription.
Hos däggdjur komprimeras ribosomalt DNA i celler eller tystas. Denna organisation verkar vara viktig för regleringen av ribosomalt DNA och för skyddet av genomisk stabilitet..
I denna region (NOR) grupperas generna (ribosomalt DNA) som kodar för ribosomalt RNA.
Kromosomerna som utgör dessa regioner varierar beroende på vilken art som studeras. Hos människor finns de i satellitregionerna i de akcentriska kromosomerna (centromeren är belägen nära en av ändarna), specifikt i par 13, 14, 15, 21 och 22.
Ribosomala DNA-enheter består av den transkriberade sekvensen och en extern distans som krävs för transkription av RNA-polymeras I.
I promotorer för ribosomalt DNA kan man skilja på två element: ett centralt element och ett uppströms element (uppströms)
Nukleolen kan betraktas som en fabrik med alla komponenter som är nödvändiga för biosyntes av ribosomprekursorer.
Ribosomalt eller ribosomalt RNA (ribonukleinsyra), vanligtvis förkortat som rRNA, är en komponent i ribosomer och deltar i proteinsyntes. Denna komponent är viktig för alla levande varelser.
Ribosomalt RNA associerar med andra proteinkomponenter. Denna bindning resulterar i ribosomala förenheter. Den ribosomala RNA-klassificeringen ges i allmänhet åtföljd av bokstaven "S", som anger Svedberg-enheterna eller sedimentationskoefficienten..
Ribosomer består av två underenheter: den stora eller stora och den lilla eller mindre..
Ribosomalt RNA från prokaryoter och eukaryoter är differentierbart. I prokaryoter är den stora subenheten 50S och består av 5S och 23S ribosomala RNA, likaså är den lilla subenheten 30S och består endast av 16S ribosomalt RNA.
Däremot består den huvudsakliga subenheten (60S) av 5S, 5.8S och 28S ribosomala RNA. Den lilla underenheten (40S) består uteslutande av 18S ribosomalt RNA.
I kärnan finns generna som kodar för ribosomala RNA: er 5.8S, 18S och 28S. Dessa ribosomala RNA transkriberas som en enda enhet i kärnan av RNA-polymeras I. Denna process resulterar i en 45S RNA-föregångare..
Nämnda ribosomala RNA-föregångare (45S) måste klyvas i dess 18S-komponenter, som tillhör den lilla underenheten (40S) och till 5.8S och 28S för den stora underenheten (60S)..
Det saknade ribosomala RNA, 5S, syntetiseras utanför kärnan; Till skillnad från motsvarigheterna katalyseras processen av RNA-polymeras III.
En cell behöver ett stort antal ribosomala RNA-molekyler. Det finns flera kopior av generna som kodar för denna typ av RNA för att uppfylla dessa höga krav.
Till exempel, baserat på data som finns i det mänskliga genomet, finns det 200 kopior för 5,8S, 18S och 28S ribosomala RNA. För 5S ribosomalt RNA finns 2000 kopior.
Processen börjar med 45S ribosomalt RNA. Det börjar med avlägsnandet av distansen nära 5'-änden. När transkriptionsprocessen är klar tas den återstående distansen placerad i 3'-änden bort. Efter efterföljande strykningar erhålls moget ribosomalt RNA.
Dessutom kräver bearbetning av ribosomalt RNA en serie viktiga modifieringar i dess baser, såsom processer för metylering och omvandling av uridin till pseudouridin..
Därefter inträffar tillsatsen av proteiner och RNA i kärnan. Dessa inkluderar små nukleolära RNA (pRNA), som deltar i separationen av ribosomala RNA i 18S-, 5.8S- och 28S-produkterna..
PRNA har sekvenser komplementära till 18S och 28S ribosomala RNA. Därför kan de modifiera baserna i föregångaren RNA, metylera vissa regioner och delta i bildandet av pseudouridin..
Ribosombildning involverar bindning av föräldraribosomalt RNA, tillsammans med ribosomala proteiner och 5S. Proteinerna som är involverade i processen transkriberas av RNA-polymeras II i cytoplasman och måste transporteras till kärnan..
Ribosomala proteiner börjar associera med ribosomala RNA innan klyvning av 45S ribosomalt RNA inträffar. Efter separation tillsätts de återstående ribosomala proteinerna och 5S ribosomalt RNA..
18S-mognad i ribosomalt RNA sker snabbare. Slutligen exporteras de "pre-ribosomala partiklarna" till cytoplasman..
Förutom biogenes av ribosomer har ny forskning visat att nucleolus är en multifunktionell enhet.
Nukleolus är också involverad i bearbetning och mognad av andra typer av RNA, såsom snRNP (protein-RNA-komplex som kombineras med pre-messenger-RNA för att bilda spliceosome- eller splice-komplex) och vissa överförings-RNA., MicroRNA och andra ribonukleoproteinkomplex.
Genom analysen av nucleolus-proteomen har proteiner associerade med RNA-bearbetning före messenger, cellcykelkontroll, DNA-replikering och reparation hittats. Proteinkonstruktionen i kärnan är dynamisk och förändras under olika miljöförhållanden och cellstress..
På samma sätt finns det en serie patologier associerade med att kärnan fungerar felaktigt. Dessa inkluderar Diamond-Blackfan anemi och neurodegenerativa störningar såsom Alzheimers och Huntingtons sjukdom..
Hos Alzheimers patienter sker en förändring av uttrycksnivåerna i nucleolus jämfört med friska patienter.
Mer än 5000 studier har visat förhållandet mellan malign cellproliferation och nucleolus-aktivitet.
Målet med vissa undersökningar är att kvantifiera nukleolproteiner för kliniska diagnostiska ändamål. Med andra ord är målet att utvärdera cancerproliferation med användning av dessa proteiner som en markör, speciellt B23-, nukleolin-, UBF- och RNA-polymeras I-underenheter..
Å andra sidan har det visat sig att B23-proteinet är direkt relaterat till utvecklingen av cancer. Likaså är andra nukleolära komponenter involverade i utvecklingen av patologier, såsom akut promyelocytisk leukemi.
Det finns tillräckliga bevis för att bekräfta att virus, både växter och djur, behöver nukleolproteiner för att uppnå replikationsprocessen. Det finns förändringar i kärnan, när det gäller morfologi och proteinkomposition, när cellen upplever en virusinfektion.
Ett betydande antal proteiner har hittats som kommer från DNA- och RNA-sekvenser som innehåller virus och är belägna i kärnan.
Virus har olika strategier som gör det möjligt för dem att lokalisera i denna subnukleära region, såsom virusproteiner som innehåller "signaler" som leder dem till kärnan. Dessa taggar är rika på aminosyrorna arginin och lysin..
Platsen för virus i kärnan underlättar deras replikering och dessutom verkar det vara ett krav för deras patogenicitet..



Ingen har kommenterat den här artikeln än.