De G-proteiner eller guaninnukleotidbindande proteiner, är proteiner associerade med plasmamembranet som tillhör en familj av signal "kopplings" -proteiner som har viktiga funktioner i många signaltransduktionsprocesser i eukaryota organismer.
I litteraturen beskrivs G-proteiner som binära molekylära switchar, eftersom deras biologiska aktivitet bestäms av förändringarna i deras struktur som ges av nukleotidarterna som de kan binda till: guanosinnukleotider (difosfat (BNP) och trifosfat (GTP)) ).
De aktiveras vanligtvis av receptorer från en familj av proteiner som kallas G-proteinkopplade receptorer (GPCR).G-proteinkopplade receptorer "), som tar emot en extern initial signal och omvandlar den till konformationsändringar som utlöser aktivering, vilket därefter översätts till aktivering av ett annat effektorprotein.
Vissa författare anser att de kodande generna för denna familj av proteiner utvecklats genom duplicering och divergens av en gemensam förfädersgen, vars produkt alltmer förfinades och specialiserades..
Bland den stora variationen av cellulära funktioner som dessa proteiner har är translokation av makromolekyler under proteinsyntes, transduktion av hormonella signaler och sensoriska stimuli, samt reglering av cellproliferation och differentiering..
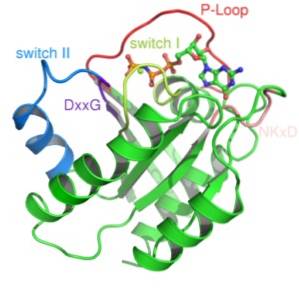
Två klasser av denna typ av protein har beskrivits: små G-proteiner och heterotrimera G-proteiner. Den första tredimensionella strukturen för ett G-protein erhölls för mer än ett decennium sedan från ett litet G-protein som kallas Ras.
Artikelindex
Strukturellt sett känns igen två typer av G-proteiner: de små G-proteinerna och de mycket mer komplexa heterotrimeriska G-proteinerna..
Små G-proteiner består av en enda polypeptid med cirka 200 aminosyrarester och cirka 20-40 kDa, och i deras struktur finns en bevarad katalytisk domän (G-domänen) som består av fem a-helixer, sex P-vikta ark och fem polypeptidöglor.
Heterotrimer G-proteiner är å andra sidan integrerade membranproteiner som består av tre polypeptidkedjor, kända som a-, P- och y-underenheter..
-A-subenheten väger mellan 40 och 52 kDa, har en guaninnukleotidbindande region och har GTPas-aktivitet för att hydrolysera bindningar mellan fosfatgrupperna i GTP..
A-underenheterna av olika G-proteiner delar vissa strukturella domäner, såsom de för bindning och hydrolys av GTP, men är mycket olika i sina bindningsställen för receptor- och effektorproteinerna..
-Β-underenheten har en något lägre molekylvikt (mellan 35 och 36 kDa).
-Y-underenheten är å andra sidan mycket mindre och har en ungefärlig molekylvikt på 8 kDa..
Alla heterotrimera G-proteiner har 7 transmembrandomäner och delar sekvenslikhet med P- och y-domänerna. Dessa två domäner är så starkt associerade att de ses som en enda funktionell enhet..
Som nämnts ovan finns det två typer av G-proteiner: små och heterotrimera..
Små G-proteiner har roller i celltillväxt, proteinsekretion och intracellulär vesikelinteraktion. För sin del är heterotrimeriska G-proteiner associerade med transduktion av signaler från ytreceptorer och fungerar också som omkopplare som växlar mellan två tillstånd beroende på den associerade nukleotiden..
Dessa proteiner kallas också små GTPaser, små GTP-bindande proteiner eller Ras-protein-superfamiljer och bildar en oberoende superfamilj inom den stora klassen GTP-hydrolaser med reglerande funktioner..
Dessa proteiner är mycket olika och kontrollerar flera cellulära processer. De kännetecknas av en bevarad GTP-bindande domän, "G" -domänen. Bindningen av denna fosfatnukleotid orsakar viktiga konformationsförändringar i dess katalytiska domän i små G-proteiner..
Dess aktivitet är nära relaterad till GTPas-aktiverande proteiner (GAP).GTPase-aktiverande protein ") och med guaninnukleotidutbytesfaktorer (GEF)Guaninnukleotidutbytesfaktor ”).
Fem klasser eller familjer av små G-proteiner har beskrivits i eukaryoter:
-Ras
-Rho
-Rab
-Sar1 / Arf
-Ran
Ras- och Rho-proteinerna kontrollerar genuttryck och Rho-proteinerna modulerar också omorganisationen av cytoskelettet. Rab- och Sar1 / Arf-gruppproteinerna påverkar vesikulär transport och Ran-proteinerna reglerar kärntransport och cellcykeln..
Denna typ av protein förtjänar också associering med två andra proteinfaktorer, så att signalvägen från den yttre miljön till det inre av cellen består av tre element i följande ordning:
Det finns en stor mångfald av heterotrimera G-proteiner och detta är relaterat till den stora mångfalden av a-underenheter som finns i naturen, där endast 20% av aminosyrasekvensen bevaras.
Vanligtvis identifieras heterotrimeriska G-proteiner tack vare a-subenhetens mångfald, baserat huvudsakligen på deras funktionella och sekvenslikheter..
Α-underenheterna består av fyra familjer (Gs-familjen, Gi / o-familjen, Gq-familjen och G12-familjen). Varje familj består av en annan ”isotyp” som tillsammans lägger till mer än 15 olika former av α-underenheter..
Denna familj innehåller representanter som också deltar i uppreglering av adenylatcyklasproteiner och uttrycks i de flesta celltyper. Den består av två medlemmar: Gs och Golf.
Prenumerationen "s " avser stimulering (från engelska "stimulering ") och prenumerationen ”olf " hänvisar till "lukt" (från engelska "Olfaction"). Golfproteiner uttrycks särskilt i sensoriska neuroner som är ansvariga för lukt.
Detta är den största och mest olika familjen. De uttrycks i många celltyper och förmedlar receptorberoende hämning av olika typer av adenylcyklas (subskriptet "jag " hänvisar till hämning).
Proteiner med a-underenheterna i Go-gruppen uttrycks särskilt i celler i centrala nervsystemet och har två varianter: A och B..
Proteiner med denna α-underenhetsfamilj är ansvariga för regleringen av fosfolipas C. Denna familj består av fyra medlemmar vars α-underenheter uttrycks av olika gener. De finns rikligt i leverceller, njurceller och lungor.
Denna familj uttrycks allmänt i organismer och det är inte känt med säkerhet vad exakt är de cellulära processerna som regleras genom proteiner med dessa underenheter..
Β- och γ-underenheter
Även om mångfalden av alfastrukturer är avgörande för identifiering av heterotrimeriska proteiner, finns det också mycket mångfald med avseende på de andra två underenheterna: beta och gamma..
G-proteiner deltar i "kanalisering" av signaler från receptorer på plasmamembranet till kanaler eller effektorenzymer..
Det vanligaste exemplet på funktionen av denna typ av protein är i regleringen av enzymet adenylatcyklas, ett enzym som ansvarar för syntesen av adenosin 3 ', 5'-monofosfat eller helt enkelt cykliskt AMP, en molekyl som har viktiga funktioner som andra budbäraren i många kända cellulära processer:
-Selektiv fosforylering av proteiner med specifika funktioner
-Genetisk transkription
-Cytoskeletal omorganisation
-Utsöndring
-Membranavpolarisering
De deltar också indirekt i regleringen av inositolsignalkaskaden (fosfatidylinositol och dess fosforylerade derivat), som är ansvariga för kontrollen av kalciumberoende processer som kemotaxi och utsöndringen av lösliga faktorer..
Många jonkanaler och transportproteiner styrs direkt av proteiner från G-proteinfamiljen, och på samma sätt är dessa proteiner involverade i många sensoriska processer som syn, lukt, bland andra..
Samspelet för ett G-protein med effektorproteinerna är specifikt för varje klass eller familj av proteiner.
För G-proteiner kopplade med membranreceptorer (heterotrimeriska G-proteiner) orsakar bindningen av en guaninnukleotid såsom BNP eller guanosindifosfat till a-underenheten associering av de tre underenheterna och bildar ett komplex som kallas Gαβγ eller G-BNP, vilket är fäst vid membranet.
Om BNP-molekylen därefter byts mot en GTP-molekyl, dissocieras den α-underenhet som är fäst till GTP från β- och γ-underenheterna och bildar ett separat komplex som kallas Gα-GTP, vilket kan förändra aktiviteten hos dess enzymer eller målbärarproteiner.
Den hydrolytiska aktiviteten hos denna underenhet gör att den kan avsluta aktiveringen, byta ut GTP mot en ny BNP och övergå till den inaktiva konformationen..
I frånvaro av de exciterade receptorerna som associeras med G-proteiner, är denna process för utbyte av BNP till GTP mycket långsam, vilket innebär att heterotrimera G-proteiner endast byter BNP mot GTP i en fysiologiskt signifikant takt när de är bundna till sina exciterade receptorer.



Ingen har kommenterat den här artikeln än.