
Två gener är kopplade när de tenderar att ärvas tillsammans som om de vore en enda enhet. Detta kan också hända med mer än två gener. I vilket fall som helst är detta beteende hos gener det som har möjliggjort genetisk kartläggning genom koppling och rekombination..
På Mendels tid hade andra forskare, såsom Boveri-makarna, observerat att det fanns kroppar i cellens kärna som utsöndrades under celldelningsprocessen. Dessa var kromosomerna.

Senare, med Morgan och hans grupps arbete, fanns det en tydligare förståelse för arv av gener och kromosomer. Det vill säga, gener separerar sig som de kromosomer som bär dem (kromosomal arvsteori).
Artikelindex
Det finns, som vi vet, mycket färre kromosomer än gener. Människan har till exempel cirka 20 000 gener fördelade i cirka 23 olika kromosomer (artens haploida belastning).
Varje kromosom representeras av en lång DNA-molekyl där många, många gener kodas separat. Varje gen befinner sig sedan vid ett visst ställe (locus) på en specifik kromosom; i sin tur bär varje kromosom många gener.
Med andra ord är alla gener på en kromosom kopplade till varandra. Om det verkar som om de inte är det beror det på att det finns en process med fysiskt utbyte av DNA mellan kromosomerna som skapar en illusion av oberoende distribution..
Denna process kallas rekombination. Om två gener är länkade men brett separerade från varandra kommer rekombination alltid att ske och generna kommer att segregera precis som Mendel observerade..
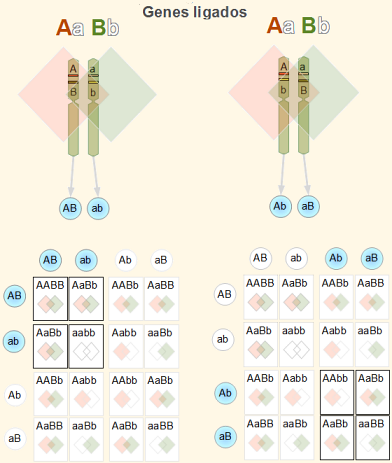
För att observera och demonstrera koppling fortsätter forskaren att göra korsningar med individer som presenterar en kontrasterande manifestation av fenotypen för generna som studeras (till exempel P: AAbb X aaBB).
Alla F1 ättlingar kommer att vara AaBb. Av dihybridkorset AaBb X aabb (eller testkors) man kan förvänta sig att en F2-avkomma visar de genotypiska (och fenotypiska) proportionerna 1AaBb:1Aabb: 1aaBb: 1aabb.
Men detta är bara sant om generna inte är länkade. Den första genetiska indikationen att två gener är kopplade är att en övervägande av faderliga fenotyper observeras: det vill säga, Aabb + aaBb >> AaB_b + aabb.
När det gäller länkade gener som vi använder som ett exempel kommer individer att producera mestadels könsceller Ab Y aB, mer än könsceller AB Y ab.
Eftersom den dominerande allelen i en gen är associerad med den recessiva allelen i den andra genen, sägs båda generna vara kopplade i avstötning. Om en övervägande av alleler observeras AB Y ab om könsceller Ab Y aB, gener sägs vara kopplade i koppling.
Det vill säga de dominerande allelerna är kopplade till samma DNA-molekyl; eller vad är detsamma, de är associerade med samma kromosom. Denna information är extremt användbar vid genetisk förbättring.
Detta gör det möjligt att fastställa antalet individer som måste analyseras när generna är länkade och det är önskvärt att välja till exempel de två dominerande tecknen.
Detta skulle vara svårare att uppnå när båda generna är i repulsion och kopplingen är så tät att det nästan inte finns någon rekombination mellan de två generna..
Förekomsten av koppling i sig var ett stort framsteg i vår förståelse av gener och deras organisation. Men dessutom tillät det oss också att förstå hur urval kan agera i befolkningar och förklara lite utvecklingen av levande varelser.
Det finns gener som är så nära kopplade att endast två typer av könsceller produceras istället för de fyra som möjliggör oberoende distribution.
I extrema fall förekommer dessa två länkade gener (i koppling eller i repulsion) bara i en typ av association i befolkningen. Om detta inträffar sägs det finnas en kopplingsjämvikt.
Länksjämvikten uppstår till exempel när bristen på de två dominerande allelerna minskar chanserna för överlevnad och reproduktion av individer..
Detta inträffar när individer är produkten av befruktning mellan könsceller ab. Befruktning mellan könsceller aB Y Ab, tvärtom ökar det sannolikheten för individens överlevnad.
Dessa kommer att ha minst en allel TILL och en allel B, och visar motsvarande vilda associerade funktioner.
Koppling och dess obalans kan också förklara varför vissa oönskade alleler av en gen inte elimineras från befolkningen. Om de är nära kopplade (i avstötning) till dominerande alleler av en annan gen som ger fördelar för deras bärare (till exempel, aB), som är associerad med det "goda" möjliggör bestående av det "dåliga".
En viktig konsekvens av koppling är att den gör det möjligt att bestämma avståndet mellan länkade gener. Detta visade sig vara historiskt sant och ledde till genereringen av de första genetiska kartorna..
För detta var det nödvändigt att förstå att homologa kromosomer kan korsa varandra under meios i en process som kallas rekombination..
Vid rekombination produceras olika könsceller till de som en individ endast kunde producera genom segregering. Eftersom rekombinanter kan räknas är det möjligt att matematiskt uttrycka hur långt ifrån varandra en gen från en annan.
I kopplings- och rekombinationskartor räknas individer som är rekombinanta mellan ett visst genpar. Sedan beräknas dess andel i förhållande till den totala sysselsatta kartläggningspopulationen..
Enligt konvention är en procent (1%) rekombination en genetisk kartenhet (umg). Till exempel, i en kartläggningspopulation på 1000 individer, finns 200 rekombinanter bland de genetiska markörerna. TILL/till Y B/b. Därför är avståndet som skiljer dem från kromosomen 20 umg.
För närvarande kallas 1 umg (vilket är 1% rekombination) cM (centi Morgan). I ovanstående fall är avståndet mellan TILL/till Y B/b är 20 cM.
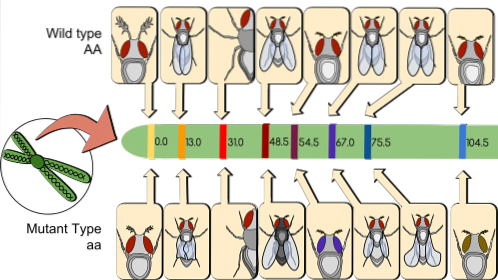
I en genetisk karta kan avstånd läggas till i cM, men uppenbarligen kan rekombinationsprocent inte läggas till. Gener bör alltid kartläggas som är tillräckligt långt ifrån varandra för att kunna mäta korta sträckor.
Om avståndet mellan två markörer är mycket högt är sannolikheten att det finns en rekombinationshändelse mellan dem lika med 1. Därför kommer de alltid att rekombineras och dessa gener kommer att bete sig som om de var oberoende fördelade trots att de är länkade..
Å andra sidan är kartorna uppmätta i cM av olika anledningar inte linjärt relaterade till mängden involverat DNA. Dessutom är mängden DNA per cM inte universell, och för varje speciell art är det ett särskilt och genomsnittligt värde..

Ingen har kommenterat den här artikeln än.