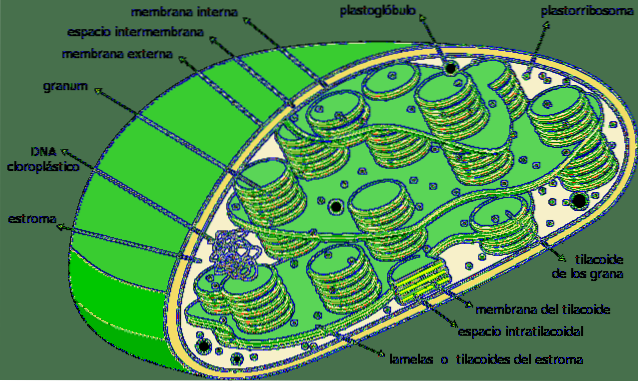
De thylakoids är fack i form av platta säckar placerade i kloroplaster i växtceller hos växter, i cyanobakterier och alger. De är vanligtvis organiserade i en struktur som kallas grana-plural granum- och det ser ut som en hög med mynt.
Thylakoids anses vara det tredje membransystemet av kloroplaster, bortsett från det inre och yttre membranet av nämnda organell. Membranet i denna struktur skiljer det inre av tylakoid från kloroplaststroma och har en serie pigment och proteiner som är involverade i metaboliska vägar..
I tylakoids finns det biokemiska reaktioner som är väsentliga för fotosyntes, en process genom vilken växter tar solljus och förvandlar det till kolhydrater. Specifikt har de nödvändiga maskinerna förankrade i sitt membran för att utföra den solberoende fasen, där ljuset fångas in och omvandlas till energi (ATP) och NADPH..
Artikelindex
Thylakoids är ett inre tredimensionellt membranöst system av kloroplaster. Helt mogna kloroplaster har 40 till 60 staplade korn, med en diameter mellan 0,3 och 0,6 µm..
Antalet tylakoider som utgör groddar varierar mycket: från färre än 10 säckar i växter som utsätts för tillräckligt med solljus till mer än 100 tylakoider i växter som lever i extremt skuggiga miljöer..
De staplade tylakoiderna är anslutna till varandra och bildar ett kontinuerligt fack i kloroplasten. Det inre av thylakoid är ett ganska rymligt fack av vattnig natur..
Tylakoidmembranet är väsentligt för fotosyntes, eftersom det första steget i processen äger rum där.
Thylakoids är de dominerande strukturerna i en helt mogen kloroplast. Om en kloroplast visualiseras i det traditionella ljusmikroskopet, kan vissa arter av korn observeras.
Dessa är de tylakoida staplarna; Av denna anledning kallade de första observatörerna av dessa strukturer dem "grana".
Med hjälp av elektronmikroskopet kunde bilden förstoras och man drog slutsatsen att naturen hos dessa korn faktiskt var staplade tylakoider.
Bildningen och strukturen för tylakoidmembranet beror på bildandet av kloroplasten från en ännu odifferentierad plastid, känd som protoplastiden. Närvaron av ljus stimulerar omvandlingen till kloroplaster och senare bildandet av staplade tylakoider.
I kloroplaster och cyanobakterier är tylakoidmembranet inte i kontakt med plasmamembranets inre del. Emellertid börjar tylakoidmembranbildningen med invagination av det inre membranet..
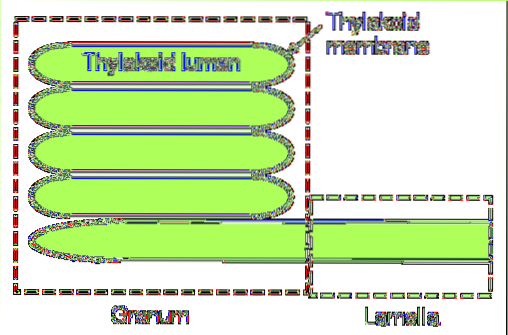
I cyanobakterier och vissa arter av alger består tylakoider av ett enda lager lameller. Däremot finns det ett mer komplext system som finns i mogna kloroplaster..
I denna sista grupp kan man skilja mellan två väsentliga delar: stromaens grana och lamell. Den första består av små staplade skivor och den andra är ansvarig för att ansluta dessa staplar med varandra och bilda en kontinuerlig struktur: tylakoidens lumen..
Lipiderna som utgör membranet är mycket specialiserade och består av nästan 80% galaktosyldiacylglycerol: monogalaktosyldiacylglycerol och digalaktosyldiacylglycerol. Dessa galaktolipider har mycket omättade kedjor, typiska för tylakoider..
På liknande sätt innehåller tylakoidmembranet mindre lipider såsom fosfatidylglycerol. De nämnda lipiderna fördelas inte homogent i båda skikten av membranet; det finns en viss grad av asymmetri som verkar bidra till att strukturen fungerar.
Fotosystem I och II är de dominerande proteinkomponenterna i detta membran. De är associerade med cytokrom b-komplexet6F- och ATP-syntetas.
Det har visat sig att de flesta elementen i fotosystem II är belägna i staplade scharlakanshinnor, medan fotosystem I mestadels är belägna i icke-staplade tylakoidmembran. Det vill säga det finns en fysisk åtskillnad mellan de två fotosystemen..
Dessa komplex innefattar integrerade membranproteiner, perifera proteiner, medfaktorer och en mängd olika pigment..
Det inre av thylakoid består av en tjock, vattnig substans vars sammansättning skiljer sig från stroma. Deltar i fotofosforylering och lagrar protonerna som genererar protonmotivkraften för syntes av ATP. I denna process kan lumenets pH nå 4.
I lumenproteomen hos modellorganismen Arabidopsis thaliana mer än 80 proteiner har identifierats, men deras funktioner har inte blivit helt klarlagda.
Lumenproteiner är involverade i regleringen av tylakoidbiogenes och i aktiviteten och omsättningen av proteiner som bildar fotosyntetiska komplex, särskilt fotosystem II och NAD (P) H-dehydrogenas..
Fotosyntesprocessen, viktig för växter, börjar i tylakoiderna. Membranet som avgränsar dem med kloroplaststroma har alla de enzymatiska maskiner som är nödvändiga för att fotosyntetiska reaktioner ska uppstå..
Fotosyntes kan delas in i två huvudfaser: ljusreaktioner och mörka reaktioner.
Som namnet antyder kan reaktioner som tillhör den första gruppen bara fortsätta i närvaro av ljus, medan de i den andra gruppen kan uppstå med eller utan ljus. Observera att det inte är nödvändigt att miljön är "mörk", den är bara oberoende av ljuset.
Den första gruppen av reaktioner, de "lätta", förekommer i tylakoid och kan sammanfattas enligt följande: ljus + klorofyll + 12 HtvåO + 12 NADP+ + 18 ADP + 18 Pi à 6 Otvå + 12 NADPH + 18 ATP.
Den andra gruppen av reaktioner förekommer i kloroplaststroma och tar upp ATP och NADPH syntetiserade i det första steget för att minska kol från koldioxid till glukos (C6H12ELLER6). Det andra steget kan sammanfattas som: 12 NADPH + 18 ATP + 6 COtvå à C6H12ELLER6 + 12 NADP+ + 18 ADP + 18 Pi + 6 timmartvåELLER.
Ljusreaktioner involverar en serie strukturer som kallas fotosystem, som finns i tylakoidmembranet och innehåller cirka 300 pigmentmolekyler, inklusive klorofyll..
Det finns två typer av fotosystem: den första har en maximal ljusabsorptionstopp på 700 nanometer och är känd som P700, medan den andra kallas P680. Båda är inbäddade i tylakoidmembranet.
Processen börjar när ett av pigmenten absorberar en foton och detta "studsar" mot andra pigment. När en klorofyllmolekyl absorberar ljus hoppar en elektron ut och en annan molekyl absorberar det. Molekylen som förlorade elektronen är nu oxiderad och har en negativ laddning.
Sidan680 fångar upp ljusenergi från klorofyll a. I detta fotosystem kastas en elektron i ett högre energisystem än en primär elektronacceptor..
Denna elektron faller in i fotosystem I och passerar genom elektrontransportkedjan. Detta system av oxidations- och reduktionsreaktioner är ansvarigt för att överföra protoner och elektroner från en molekyl till en annan..
Med andra ord finns det ett flöde av elektroner från vatten till fotosystem II, fotosystem I och NADPH..
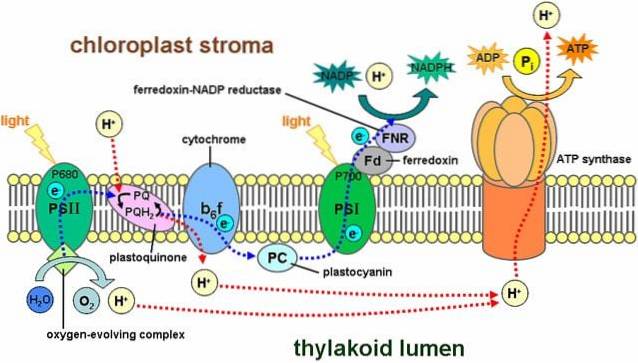
En del av protonerna som genereras av detta reaktionssystem är placerat inuti tylakoid (även kallat thylakoidljus), vilket skapar en kemisk gradient som genererar en protonmotivkraft..
Protoner rör sig från tylakoidutrymmet till stroma och följer den elektrokemiska gradienten fördelaktigt; det vill säga de kommer ut ur tylakoid.
Protonernas passage går dock inte genom något ställe i membranet, de måste göra det genom ett komplext enzymatiskt system som kallas ATP-syntetas.
Denna rörelse av protoner mot stroma orsakar bildandet av ATP med start från ADP, en process som är analog med den som sker i mitokondrierna. Syntesen av ATP med hjälp av ljus kallas fotofosforylering.
Dessa nämnda steg inträffar samtidigt: klorofyllen i fotosystem II förlorar en elektron och måste ersätta den med en elektron från nedbrytningen av en vattenmolekyl; fotosystem I fångar upp ljus, oxiderar och släpper ut en elektron som fångas av NADP+.
Den förlorade elektronen från fotosystem I ersätts med den som härrör från fotosystem II. Dessa föreningar kommer att användas i efterföljande kolfixeringsreaktioner, i Calvin-cykeln..
Utvecklingen av fotosyntes som en syrefrisättande process tillät liv som vi känner det.
Det föreslås att fotosyntes utvecklades för några miljarder år sedan i förfadern som gav upphov till nuvarande cyanobakterier, från ett anoxiskt fotosyntetiskt komplex..
Det föreslås att utvecklingen av fotosyntes åtföljs av två oumbärliga händelser: skapandet av fotosystem P680 och uppkomsten av ett inre membransystem utan anslutning till cellmembranet.
Det finns ett protein som kallas Vipp1 som är viktigt för bildandet av tylakoider. Faktum är att detta protein finns i växter, alger och cyanobakterier, men frånvarande i bakterier som utför anoxisk fotosyntes..
Man tror att denna gen kunde ha sitt ursprung genom genduplicering hos den möjliga förfäder till cyanobakterier. Det finns bara ett fall av cyanobakterier som kan fotosyntes med syre och inte har tylakoider: arten Gloeobacter violaceus.


Ingen har kommenterat den här artikeln än.