De sätt av pentoser Fosfat, även känd som avledning av hexosmonofosfat, är en grundläggande metabolisk väg vars slutprodukt är riboser, nödvändig för syntesvägar för nukleotid och nukleinsyra, såsom DNA, RNA, ATP, NADH, FAD och koenzym A.
Det producerar också NADPH (nikotinamidadenindinukleotidfosfat), som används i olika enzymatiska reaktioner. Denna väg är väldigt dynamisk och kan anpassa sina produkter beroende på cellernas tillfälliga behov..

ATP (adenosintrifosfat) anses vara cellens "energivaluta", eftersom dess hydrolys kan kopplas till ett brett spektrum av biokemiska reaktioner..
På samma sätt är NADPH en viktig andra energivaluta för bland annat den reduktiva syntesen av fettsyror, kolesterolsyntes, neurotransmittorsyntes, fotosyntes och avgiftning..
Även om NADPH och NADH har samma struktur, kan de inte användas omväxlande i biokemiska reaktioner. NADPH deltar i användningen av fri energi vid oxidation av vissa metaboliter för reduktiv biosyntes.
Däremot är NADH inblandad i användningen av fri energi från oxidationen av metaboliter för att syntetisera ATP..
Artikelindex
Indikationerna om förekomsten av denna väg började 1930 tack vare forskaren Otto Warburg, som krediteras upptäckten av NADP+.
Vissa observationer tillät upptäckten av vägen, särskilt fortsättningen av andning i närvaro av glykolyshämmare, såsom fluoridjonen..
1950 beskrev forskarna Frank Dickens, Bernard Horecker, Fritz Lipmann och Efraim Racker pentosfosfatvägen.
Vävnader involverade i syntesen av kolesterol och fettsyror, såsom bröstkörtlar, fettvävnad och njurar, har höga koncentrationer av pentosfosfatenzymer..
Levern är också en viktig vävnad för denna väg: cirka 30% av glukosoxidationen i denna vävnad sker tack vare enzymerna i pentosfosfatvägen..
Pentosfosfatvägen är ansvarig för att bibehålla kolhomeostas i cellen. På samma sätt syntetiserar vägen föregångarna till nukleotider och molekyler som är involverade i syntesen av aminosyror (byggstenarna för peptider och proteiner)..
Det är den huvudsakliga källan till att minska kraften för enzymatiska reaktioner. Dessutom tillhandahåller den de molekyler som är nödvändiga för anabola reaktioner och för försvarsprocesser mot oxidativ stress. Den sista fasen av vägen är kritisk i redoxprocesser under stresssituationer.
Pentosfosfatvägen består av två faser i cellcytosolen: en oxidativ som genererar NADPH med oxidationen av glukos-6-fosfat till ribos-5-fosfat; och en icke-oxidativ, som involverar omvandlingen av sockerarter av tre, fyra, fem, sex och sju kolatomer.
Denna rutt presenterar reaktioner som delas med Calvin-cykeln och med Entner-Doudoroff-vägen, som är ett alternativ till glykolys..
Den oxidativa fasen börjar med dehydrogenering av glukos-6-fosfatmolekylen vid kol 1. Denna reaktion katalyseras av enzymet glukos-6-fosfatdehydrogenas, som har en hög specificitet för NADP+.
Produkten av denna reaktion är 6-fosfonoglukono-5-lakton. Denna produkt hydrolyseras sedan av enzymet laktonas för att ge 6-fosfoglukonat. Denna sista förening tas upp av enzymet 6-fosfoglukonatdehydrogenas och blir ribulos 5-fosfat.
Enzymet fosfopentosisomeras katalyserar det sista steget i den oxidativa fasen, vilket involverar syntesen av ribos 5-fosfat genom isomerisering av ribulos 5-fosfat.
Denna serie av reaktioner producerar två molekyler NADPH och en molekyl ribos 5-fosfat för varje molekyl glukos-6-fosfat som kommer in i denna enzymatiska väg..
I vissa celler är kraven för NADPH större än för ribos-5-fosfat. Därför tar enzymerna transketolas och transaldolas ribos 5-fosfat och omvandlar det till glyceraldehyd-3-fosfat och fruktos 6-fosfat, vilket viker för den icke-oxidativa fasen. Dessa två sista föreningar kan komma in i den glykolytiska vägen.
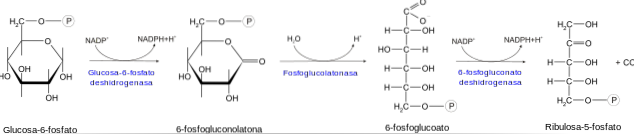
Fasen börjar med en epimeriseringsreaktion katalyserad av enzymet pentos-5-fosfatepimeras. Ribulosa-5-fosfat tas upp av detta enzym och omvandlas till xylulosa-5-fosfat.
Produkten tas upp av enzymet transketolas som verkar tillsammans med koenzymtiaminpyrofosfat (TTP), som katalyserar passage av xylulosa-5-fosfat till ribos-5-fosfat. Med överföringen av ketos till aldos produceras glyceraldehyd-3-fosfat och sedoheptulos-7-fosfat.
Enzymtransaldolas överför sedan C3 från sedoheptulos-7-fosfatmolekylen till glyceraldehyd-3-fosfat, vilket ger ett socker med fyra kol (erytros-4-fosfat) och ett sex-kolsocker (fruktos-6-fosfat). Dessa produkter kan mata den glykolytiska vägen.
Enzymet transketosala verkar igen för att överföra en C2 från xylulosa-5-fosfat till erytros-4-fosfat, vilket resulterar i fruktos-6-fosfat och glyceraldehyd-3-fosfat. Som i föregående steg kan dessa produkter gå in i glykolys.
Denna andra fas förbinder vägarna som genererar NADPH med de som ansvarar för att syntetisera ATP och NADH. Dessutom kan produkterna fruktos-6-fosfat och glyceraldehyd-3-fosfat komma in i glukoneogenes..
Olika patologier är relaterade till pentosfosfatvägen, mellan dessa neuromuskulära sjukdomar och olika typer av cancer.
De flesta kliniska studier fokuserar på att kvantifiera aktiviteten av glukos-6-fosfatdehydrogenas, eftersom det är det huvudsakliga enzymet som ansvarar för att reglera vägen.
I blodceller som tillhör individer som är mottagliga för anemi uppvisar de en låg enzymatisk aktivitet av glukos-6-fosfatdehydrogenas. Däremot uppvisar cellinjer relaterade till karcinom i struphuvudet hög enzymaktivitet..
NADPH är inblandad i produktionen av glutation, en nyckelpeptidmolekyl i skydd mot reaktiva syrearter, involverad i oxidativ stress.
Olika typer av cancer leder till aktivering av pentosvägen och den är associerad med metastaseringsprocesser, angiogenes och svar på kemoterapi och strålbehandling..
Å andra sidan utvecklas kronisk granulomatös sjukdom när det finns en brist i produktionen av NADPH.



Ingen har kommenterat den här artikeln än.